

Cardiovascular consequences of KATP overactivity in Cantu syndrome
- PMID: 30089727
- PMCID: PMC6129117
- DOI: 10.1172/jci.insight.121153
Cardiovascular consequences of KATP overactivity in Cantu syndrome
Abstract
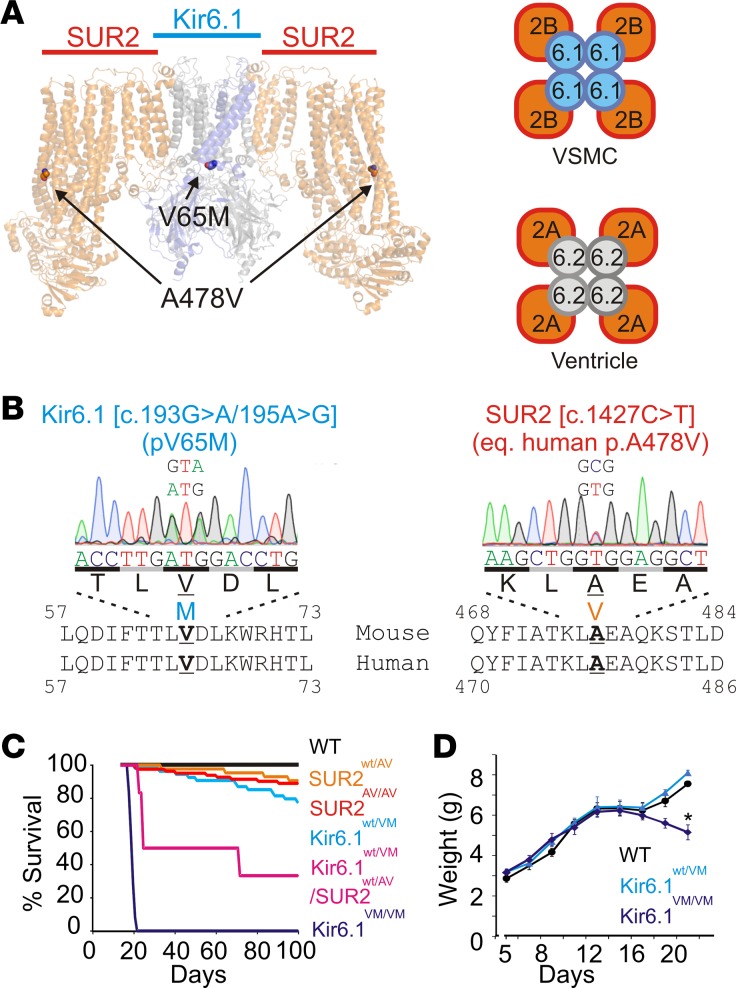
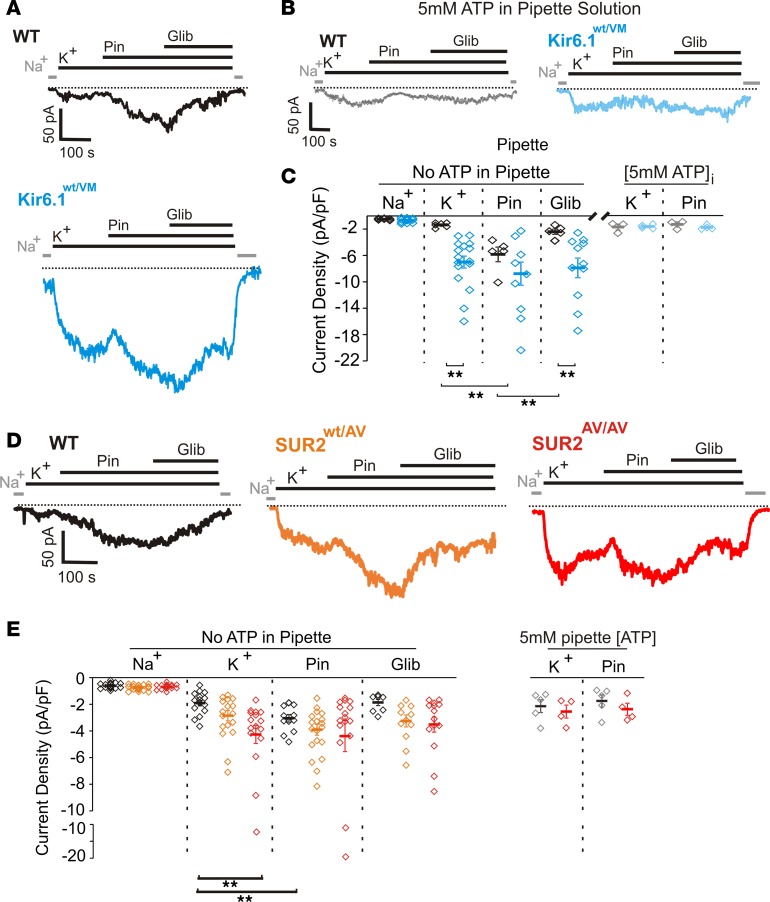
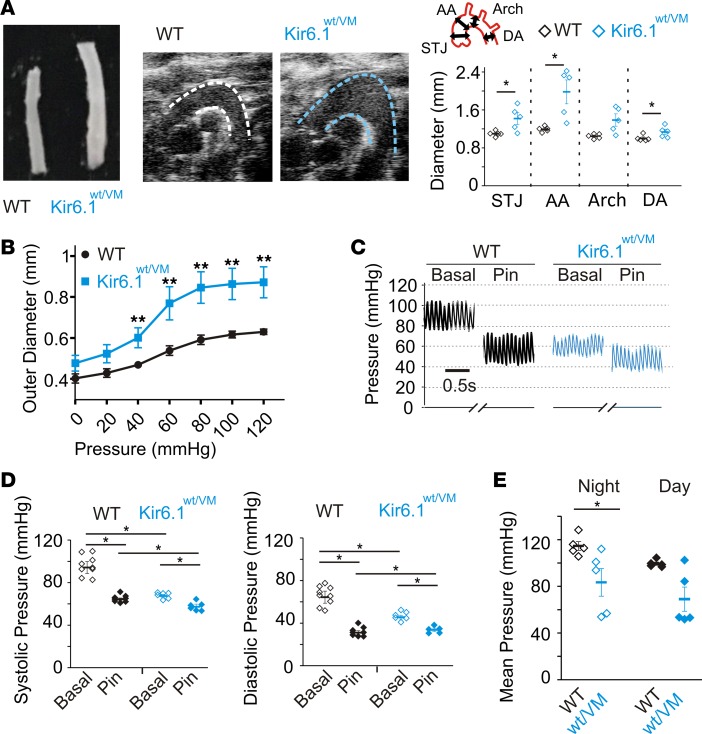
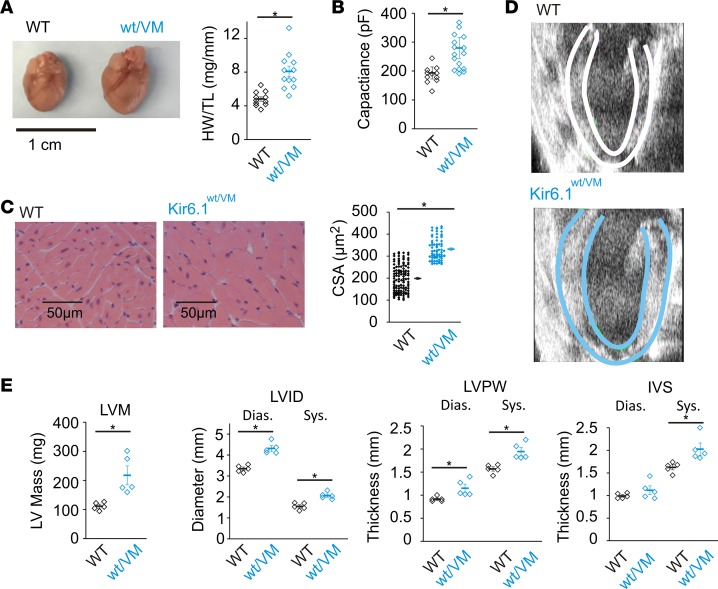
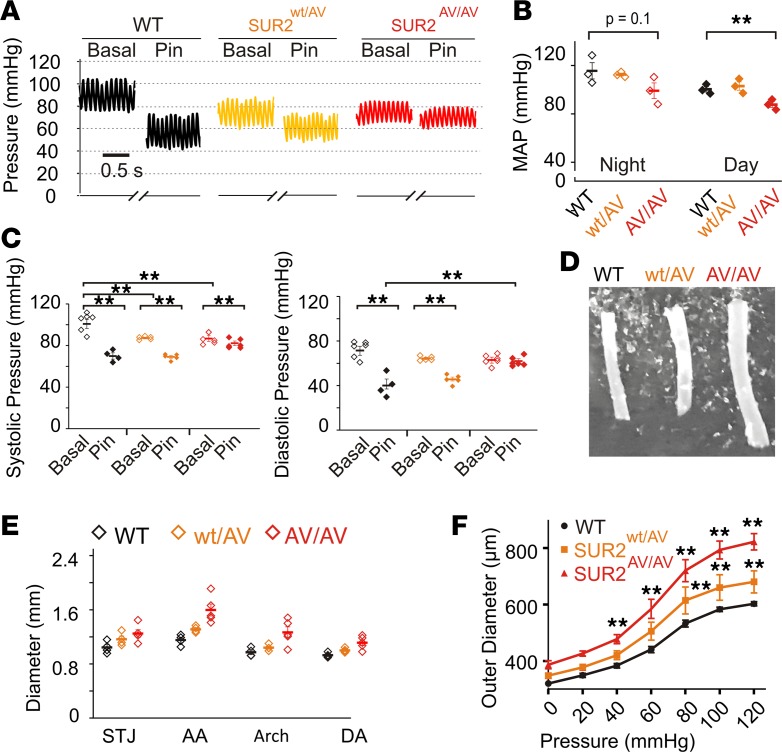
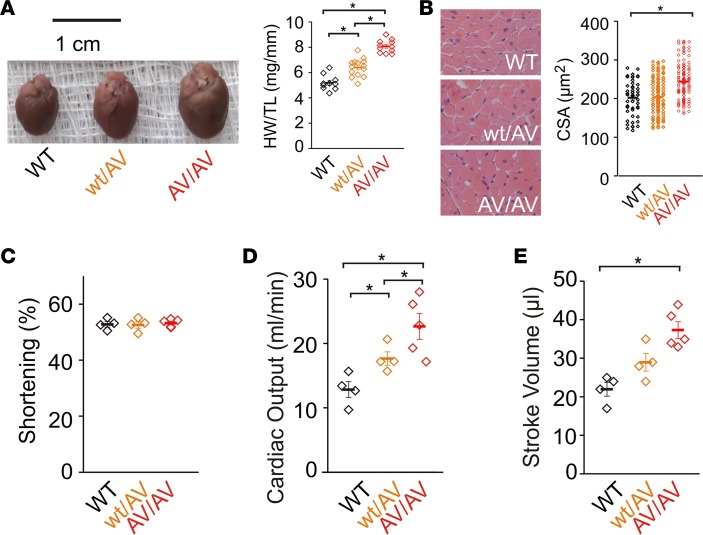
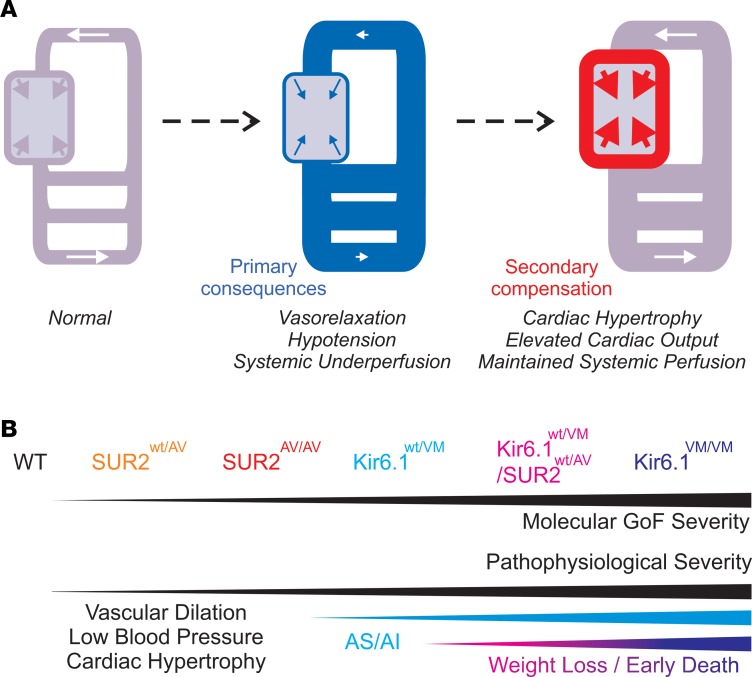
Cantu syndrome (CS) is characterized by multiple vascular and cardiac abnormalities including vascular dilation and tortuosity, systemic hypotension, and cardiomegaly. The disorder is caused by gain-of-function (GOF) mutations in genes encoding pore-forming (Kir6.1, KCNJ8) and accessory (SUR2, ABCC9) ATP-sensitive potassium (KATP) channel subunits. However, there is little understanding of the link between molecular dysfunction and the complex pathophysiology observed, and there is no known treatment, in large part due to the lack of appropriate preclinical disease models in which to test therapies. Notably, expression of Kir6.1 and SUR2 does not fully overlap, and the relative contribution of KATP GOF in various cardiovascular tissues remains to be elucidated. To investigate pathophysiologic mechanisms in CS we have used CRISPR/Cas9 engineering to introduce CS-associated SUR2[A478V] and Kir6.1[V65M] mutations to the equivalent endogenous loci in mice. Mirroring human CS, both of these animals exhibit low systemic blood pressure and dilated, compliant blood vessels, as well dramatic cardiac enlargement, the effects being more severe in V65M animals than in A478V animals. In both animals, whole-cell patch-clamp recordings reveal enhanced basal KATP conductance in vascular smooth muscle, explaining vasodilation and lower blood pressure, and demonstrating a cardinal role for smooth muscle KATP dysfunction in CS etiology. Echocardiography confirms in situ cardiac enlargement and increased cardiac output in both animals. Patch-clamp recordings reveal reduced ATP sensitivity of ventricular myocyte KATP channels in A478V, but normal ATP sensitivity in V65M, suggesting that cardiac remodeling occurs secondary to KATP overactivity outside of the heart. These SUR2[A478V] and Kir6.1[V65M] animals thus reiterate the key cardiovascular features seen in human CS. They establish the molecular basis of the pathophysiological consequences of reduced smooth muscle excitability resulting from SUR2/Kir6.1-dependent KATP GOF, and provide a validated animal model in which to examine potential therapeutic approaches to treating CS.
Keywords: Cardiology; Cardiovascular disease; Genetic diseases; Ion channels.
Conflict of interest statement
Figures



References
-
- Scurr I, et al. Cantú syndrome: report of nine new cases and expansion of the clinical phenotype. Am J Med Genet A. 2011;155A(3):508–518. - PubMed
-
- Grange DK, Lorch SM, Cole PL, Singh GK. Cantu syndrome in a woman and her two daughters: Further confirmation of autosomal dominant inheritance and review of the cardiac manifestations. Am J Med Genet A. 2006;140(15):1673–1680. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
