

Precision gene editing technology and applications in nephrology
- PMID: 30089813
- PMCID: PMC6591726
- DOI: 10.1038/s41581-018-0047-x
Precision gene editing technology and applications in nephrology
Abstract
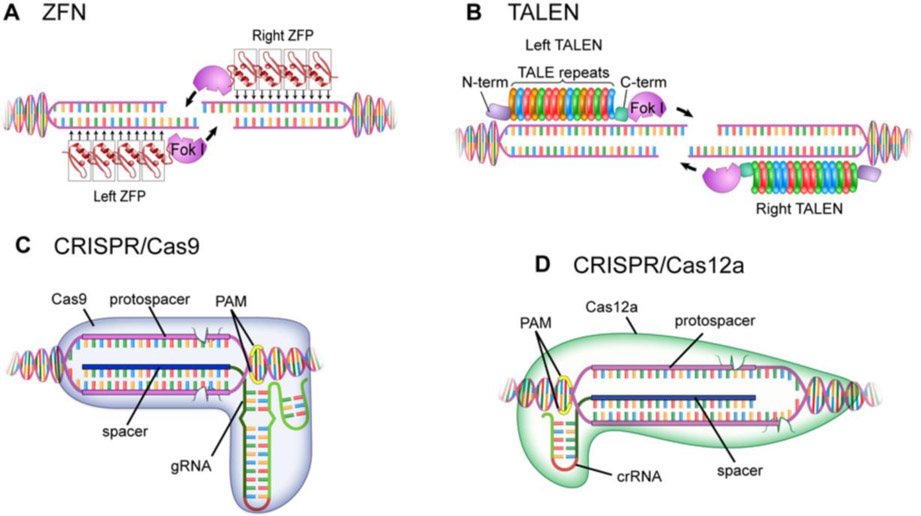
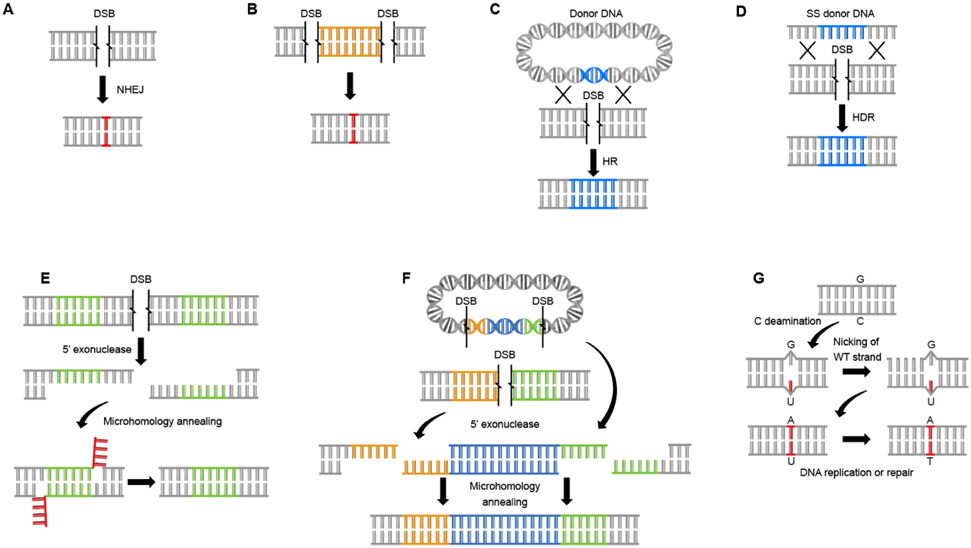
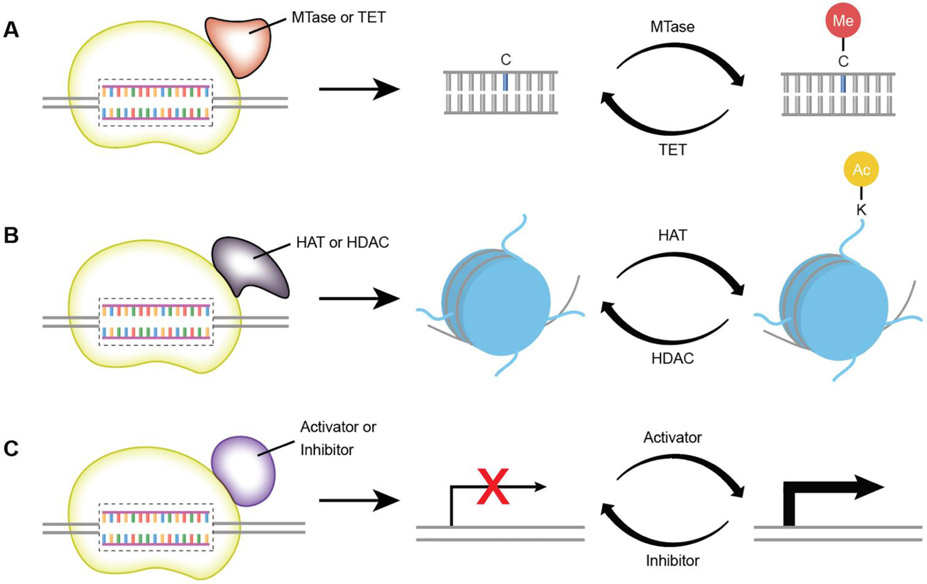
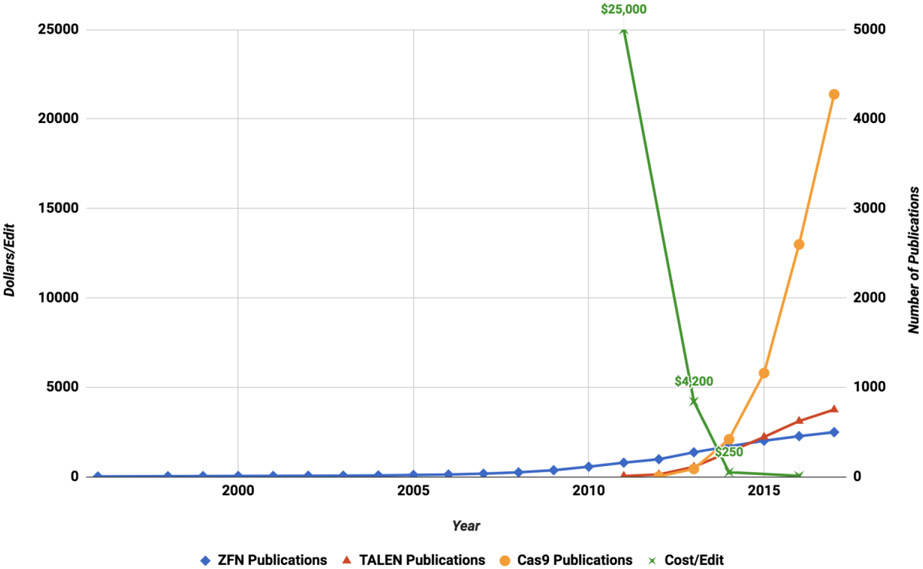
The expanding field of precision gene editing is empowering researchers to directly modify DNA. Gene editing is made possible using synonymous technologies: a DNA-binding platform to molecularly locate user-selected genomic sequences and an associated biochemical activity that serves as a functional editor. The advent of accessible DNA-targeting molecular systems, such as zinc-finger nucleases, transcription activator-like effectors (TALEs) and CRISPR-Cas9 gene editing systems, has unlocked the ability to target nearly any DNA sequence with nucleotide-level precision. Progress has also been made in harnessing endogenous DNA repair machineries, such as non-homologous end joining, homology-directed repair and microhomology-mediated end joining, to functionally manipulate genetic sequences. As understanding of how DNA damage results in deletions, insertions and modifications increases, the genome becomes more predictably mutable. DNA-binding platforms such as TALEs and CRISPR can also be used to make locus-specific epigenetic changes and to transcriptionally enhance or suppress genes. Although many challenges remain, the application of precision gene editing technology in the field of nephrology has enabled the generation of new animal models of disease as well as advances in the development of novel therapeutic approaches such as gene therapy and xenotransplantation.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
