

Immunotherapy targeting toll-like receptor 2 alleviates neurodegeneration in models of synucleinopathy by modulating α-synuclein transmission and neuroinflammation
- PMID: 30092810
- PMCID: PMC6085656
- DOI: 10.1186/s13024-018-0276-2
Immunotherapy targeting toll-like receptor 2 alleviates neurodegeneration in models of synucleinopathy by modulating α-synuclein transmission and neuroinflammation
Abstract
Background: Synucleinopathies of the aging population are an heterogeneous group of neurological disorders that includes Parkinson's disease (PD) and dementia with Lewy bodies (DLB) and are characterized by the progressive accumulation of α-synuclein in neuronal and glial cells. Toll-like receptor 2 (TLR2), a pattern recognition immune receptor, has been implicated in the pathogenesis of synucleinopathies because TLR2 is elevated in the brains of patients with PD and TLR2 is a mediator of the neurotoxic and pro-inflammatory effects of extracellular α-synuclein aggregates. Therefore, blocking TLR2 might alleviate α-synuclein pathological and functional effects. For this purpose, herein, we targeted TLR2 using a functional inhibitory antibody (anti-TLR2).
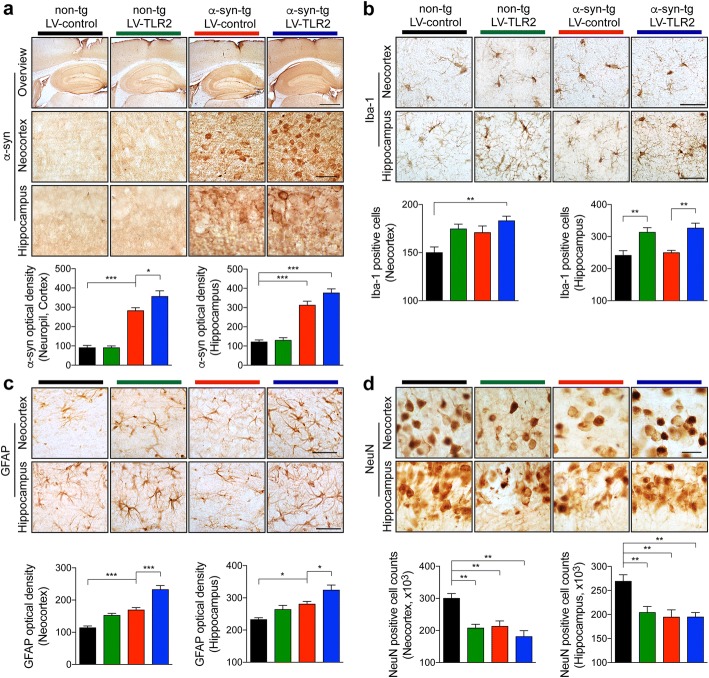
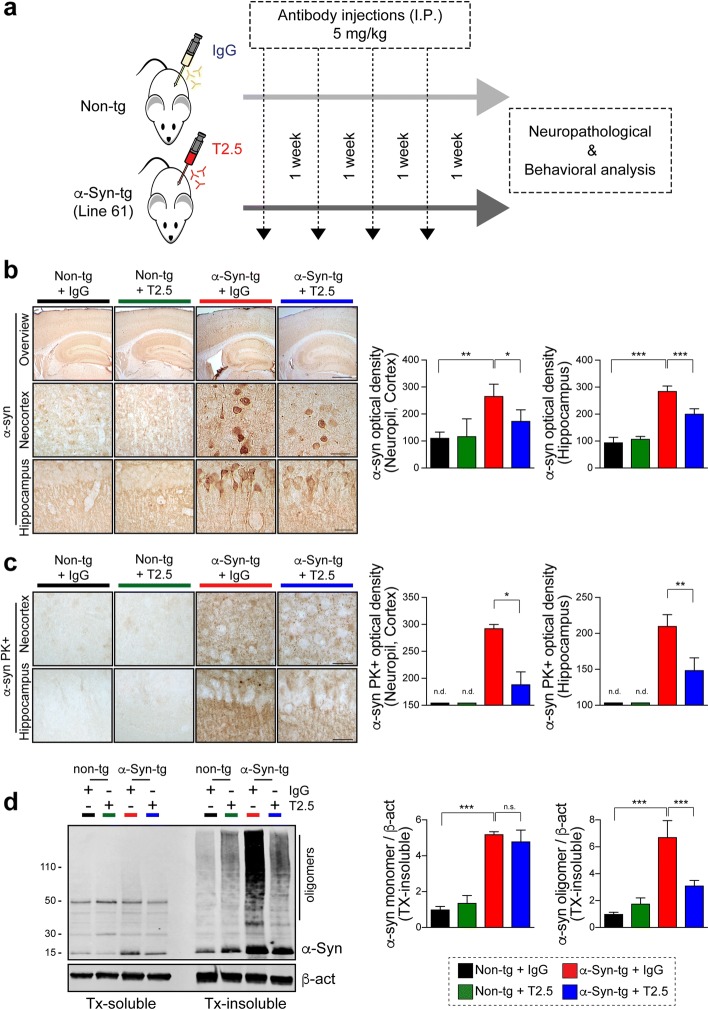
Methods: Two different human α-synuclein overexpressing transgenic mice were used in this study. α-synuclein low expresser mouse (α-syn-tg, under the PDGFβ promoter, D line) was stereotaxically injected with TLR2 overexpressing lentivirus to demonstrate that increment of TLR2 expression triggers neurotoxicity and neuroinflammation. α-synuclein high expresser mouse (α-Syn-tg; under mThy1 promoter, Line 61) was administrated with anti-TLR2 to examine that functional inhibition of TLR2 ameliorates neuropathology and behavioral defect in the synucleinopathy animal model. In vitro α-synuclein transmission live cell monitoring system was used to evaluate the role of TLR2 in α-synuclein cell-to-cell transmission.
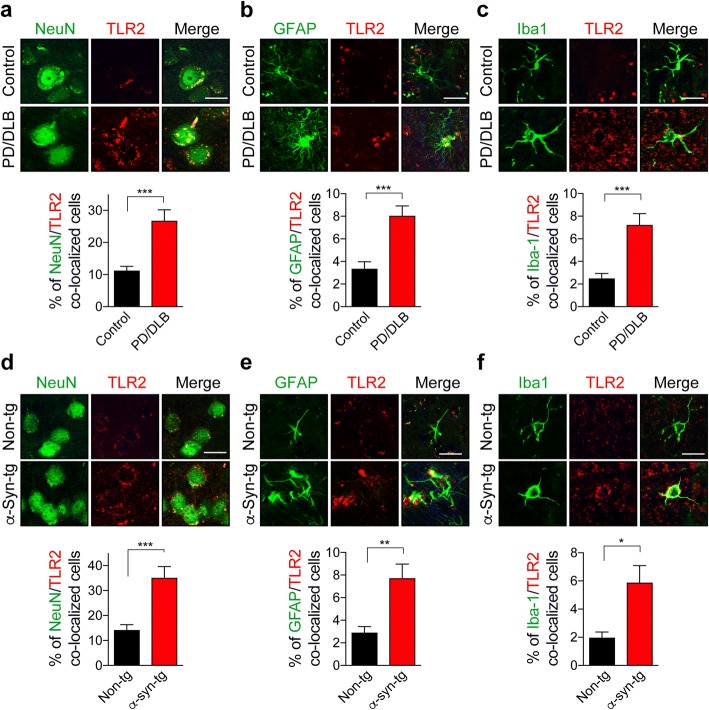
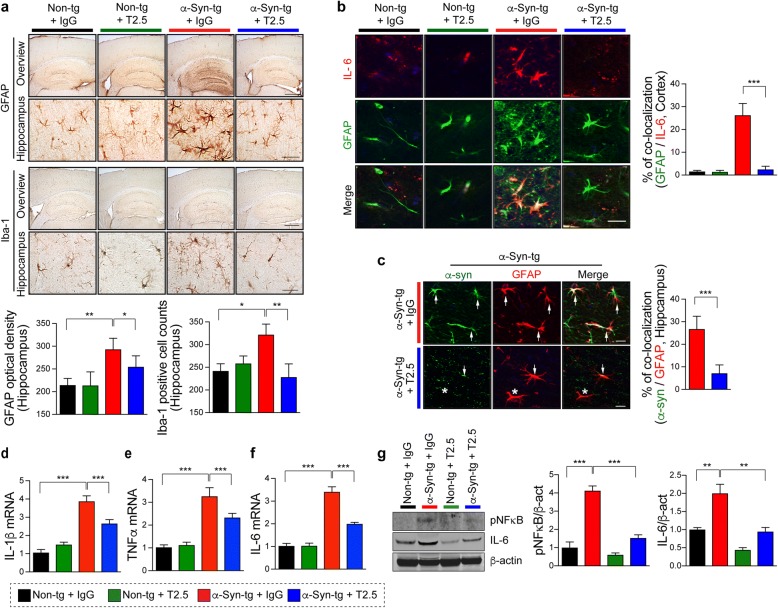
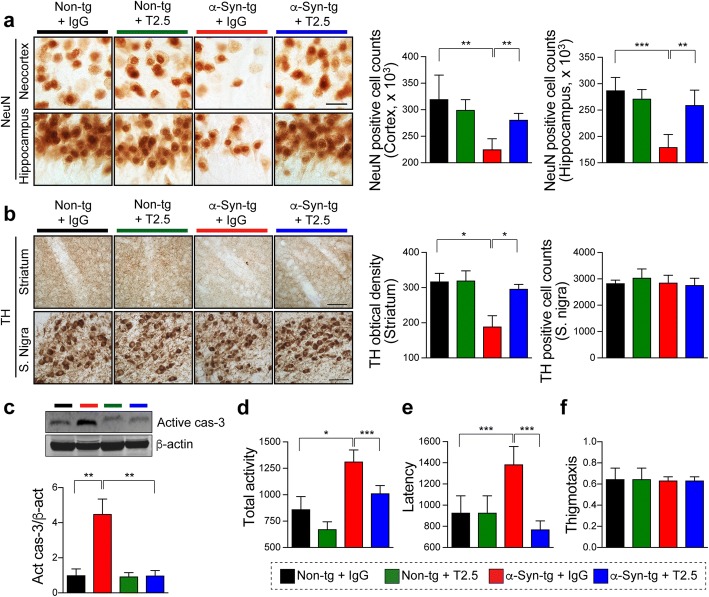
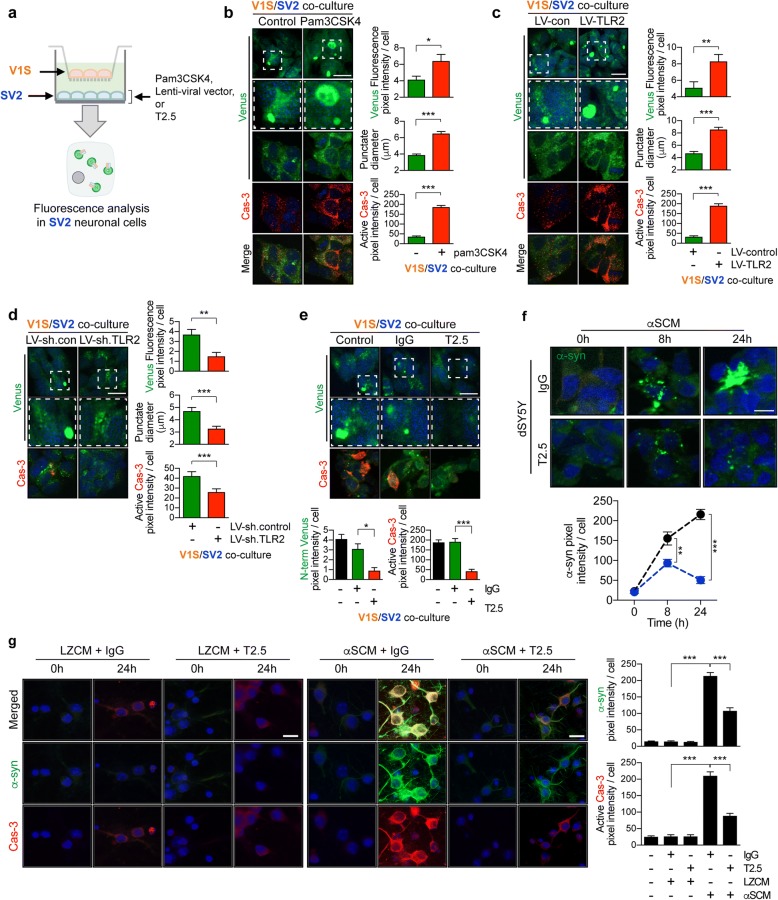
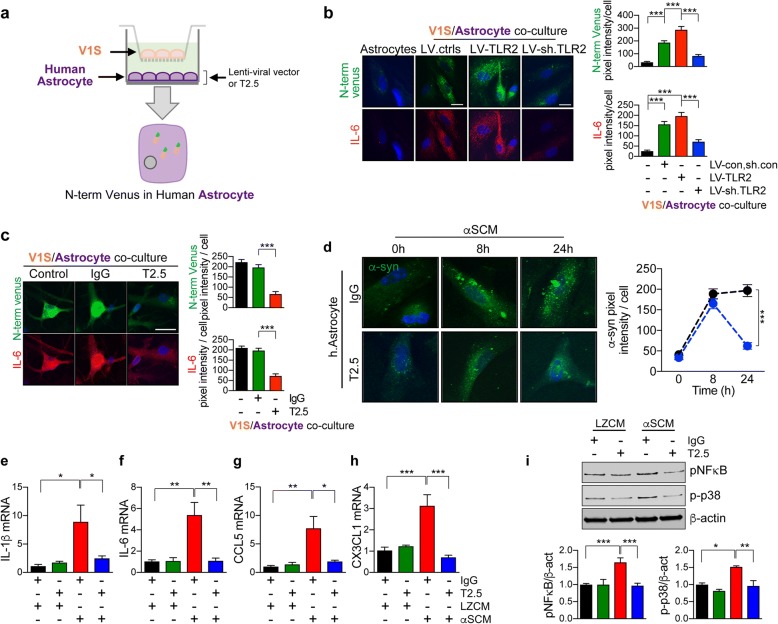
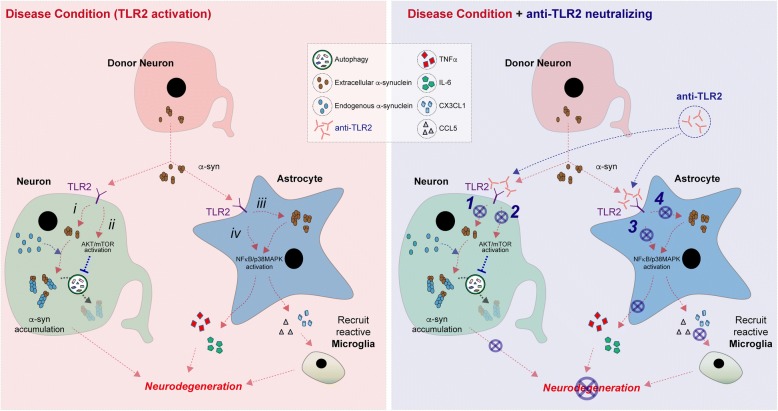
Results: We demonstrated that administration of anti-TLR2 alleviated α-synuclein accumulation in neuronal and astroglial cells, neuroinflammation, neurodegeneration, and behavioral deficits in an α-synuclein tg mouse model of PD/DLB. Moreover, in vitro studies with neuronal and astroglial cells showed that the neuroprotective effects of anti-TLR2 antibody were mediated by blocking the neuron-to-neuron and neuron-to-astrocyte α-synuclein transmission which otherwise promotes NFκB dependent pro-inflammatory responses.
Conclusion: This study proposes TLR2 immunotherapy as a novel therapeutic strategy for synucleinopathies of the aging population.
Keywords: Immunotherapy; Neurodegeneration; Neuroinflammation; Parkinson’s disease; Synucleinopathy; Toll-like receptor 2; Transmission; α-synuclein.
Conflict of interest statement
All procedures for animal use were approved by the institutional Animal Care and Use Committee at University of California, San Diego under protocol S02221.
All authors have approved of the consents of this manuscript and provided consent for publication.
The authors declare that they have no competing interests.
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
