

Apoptosis propagates through the cytoplasm as trigger waves
- PMID: 30093599
- PMCID: PMC6263143
- DOI: 10.1126/science.aah4065
Apoptosis propagates through the cytoplasm as trigger waves
Abstract
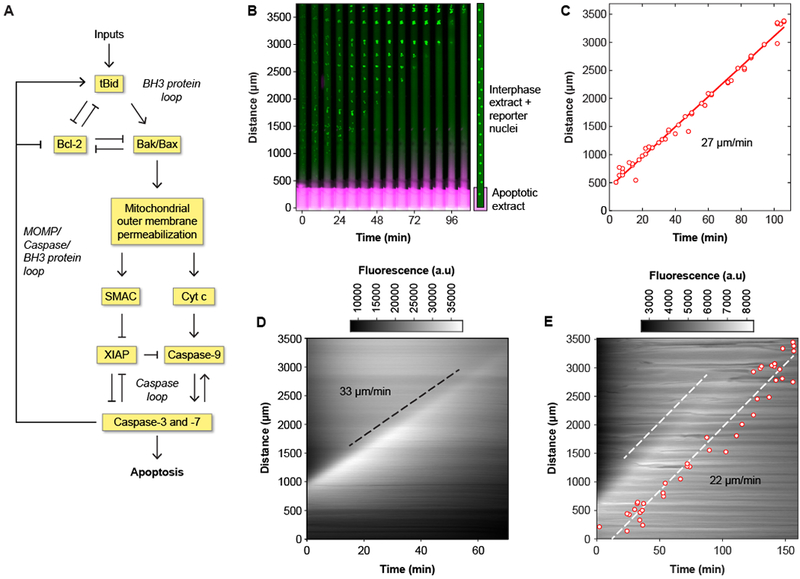
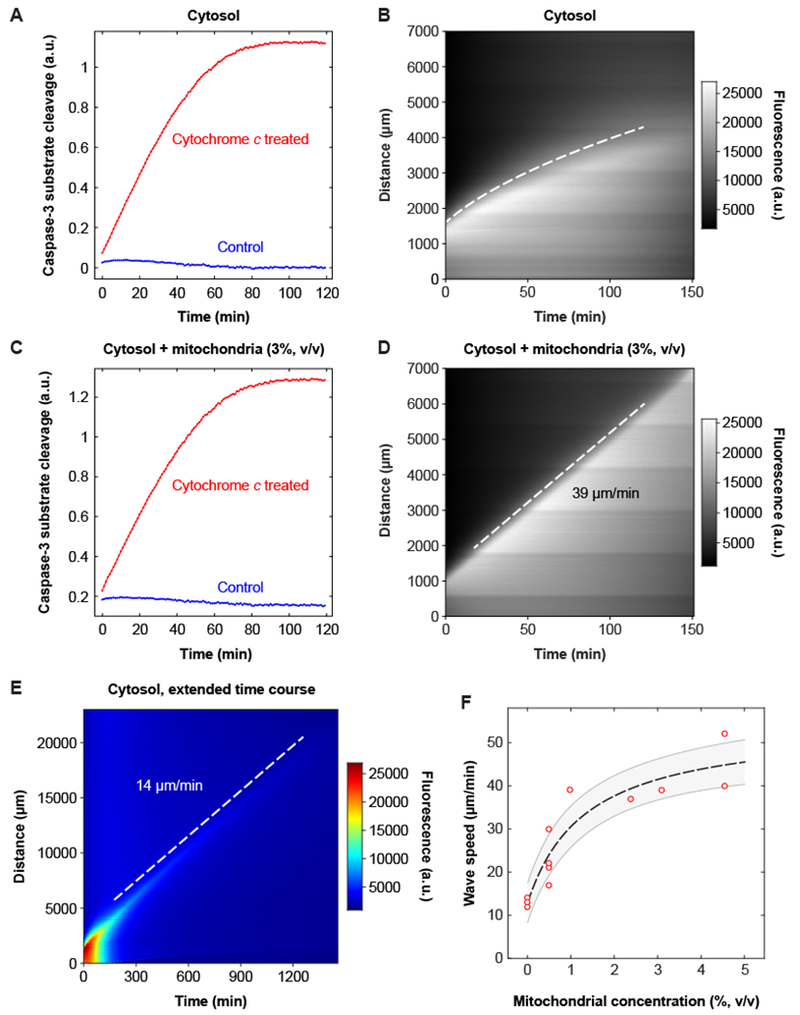
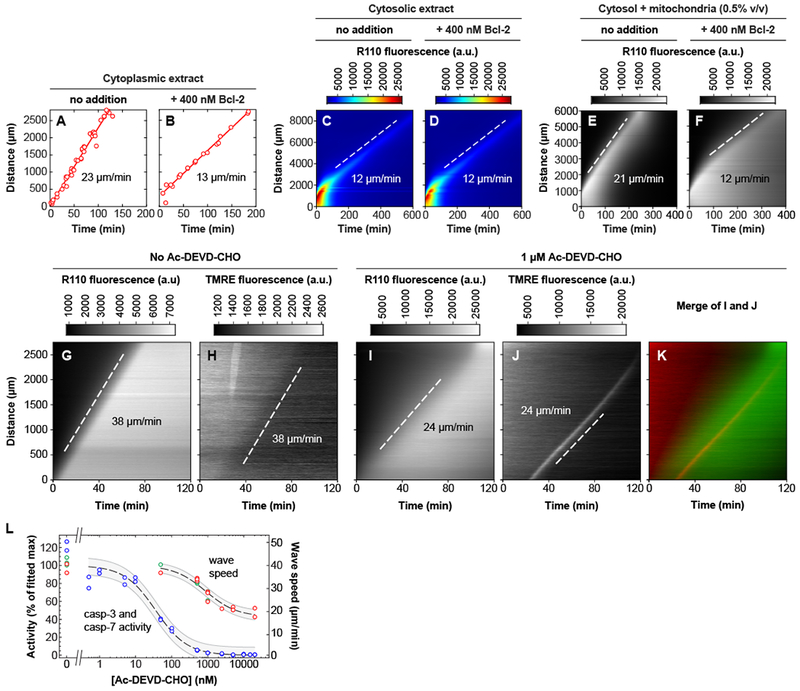
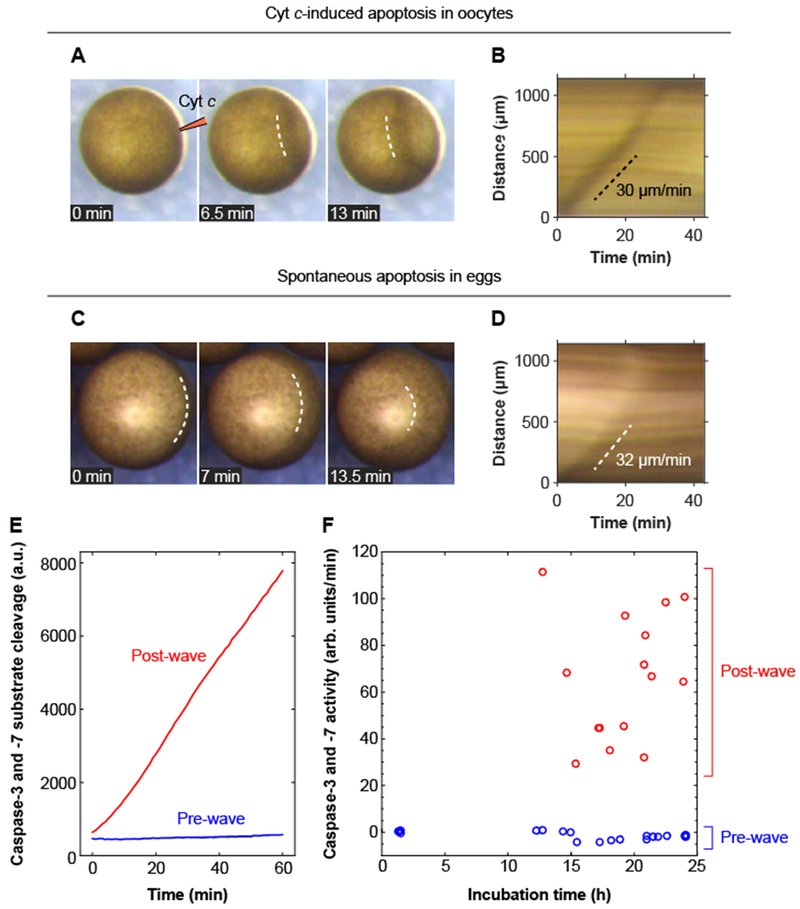
Apoptosis is an evolutionarily conserved form of programmed cell death critical for development and tissue homeostasis in animals. The apoptotic control network includes several positive feedback loops that may allow apoptosis to spread through the cytoplasm in self-regenerating trigger waves. We tested this possibility in cell-free Xenopus laevis egg extracts and observed apoptotic trigger waves with speeds of ~30 micrometers per minute. Fractionation and inhibitor studies implicated multiple feedback loops in generating the waves. Apoptotic oocytes and eggs exhibited surface waves with speeds of ~30 micrometers per minute, which were tightly correlated with caspase activation. Thus, apoptosis spreads through trigger waves in both extracts and intact cells. Our findings show how apoptosis can spread over large distances within a cell and emphasize the general importance of trigger waves in cell signaling.
Copyright © 2018, American Association for the Advancement of Science.
Conflict of interest statement
Figures
References
-
- Newmeyer DD, Farschon DM, Reed JC, Cell-free apoptosis in Xenopus egg extracts: inhibition by Bcl-2 and requirement for an organelle fraction enriched in mitochondria. Cell 79, 353–364 (1994). - PubMed
-
- Deming P, Kornbluth S, Study of apoptosis in vitro using the Xenopus egg extract reconstitution system. Meth. Mol. Biol 322, 379–393 (2006). - PubMed
-
- Tyson JJ, Keener JP, Singular perturbation theory of traveling waves in excitable media (a review). Physica D 32, 327–361 (1988).
-
- Winfree AT, Two kinds of wave in an oscillating chemical solution. Faraday Symp. Chem. Soc 9, 38–46 (1974).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
