

Developmental barcoding of whole mouse via homing CRISPR
- PMID: 30093604
- PMCID: PMC6139672
- DOI: 10.1126/science.aat9804
Developmental barcoding of whole mouse via homing CRISPR
Abstract
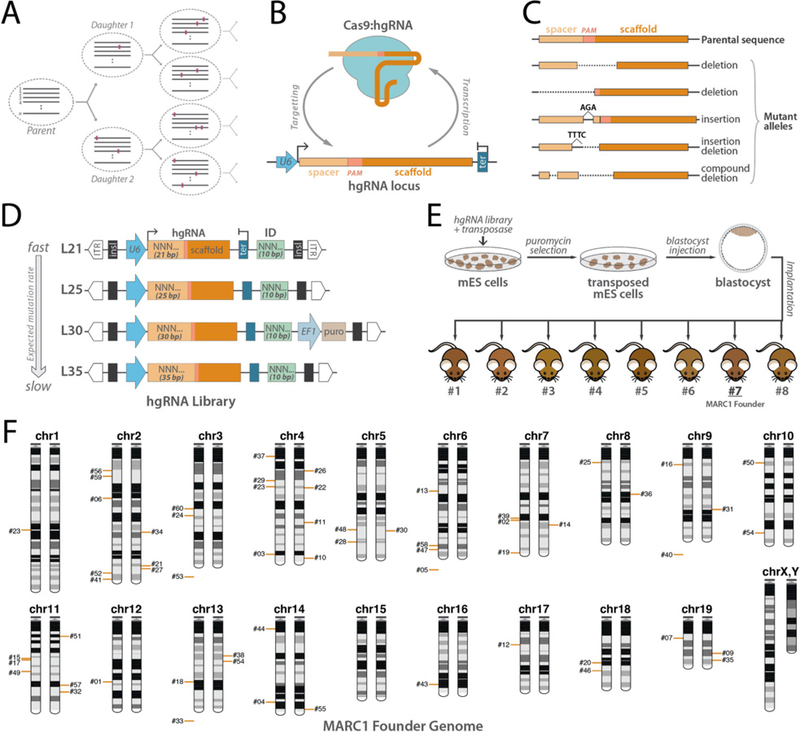
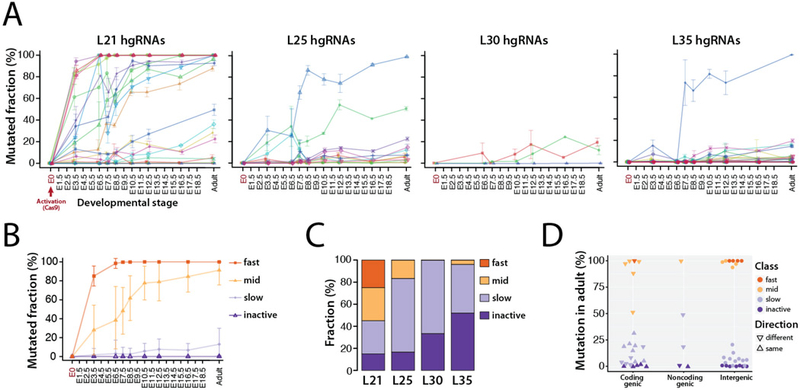
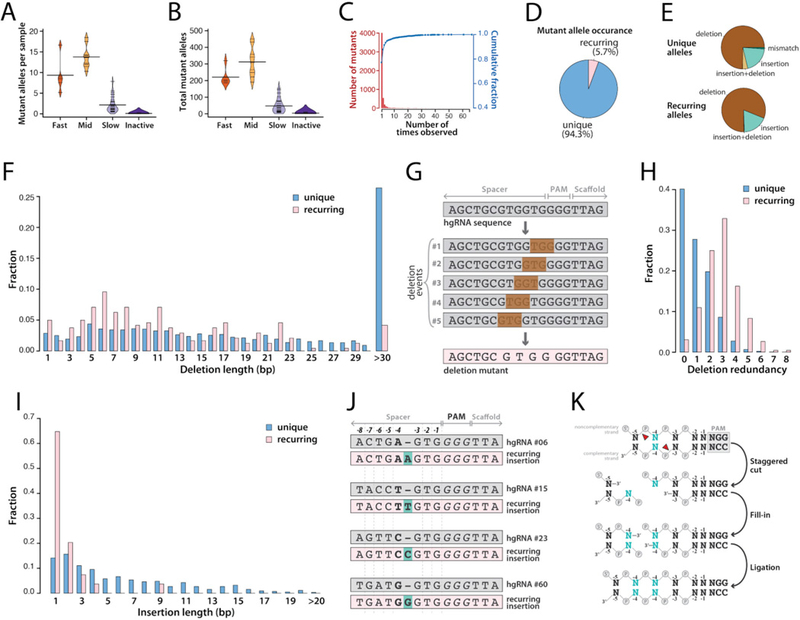
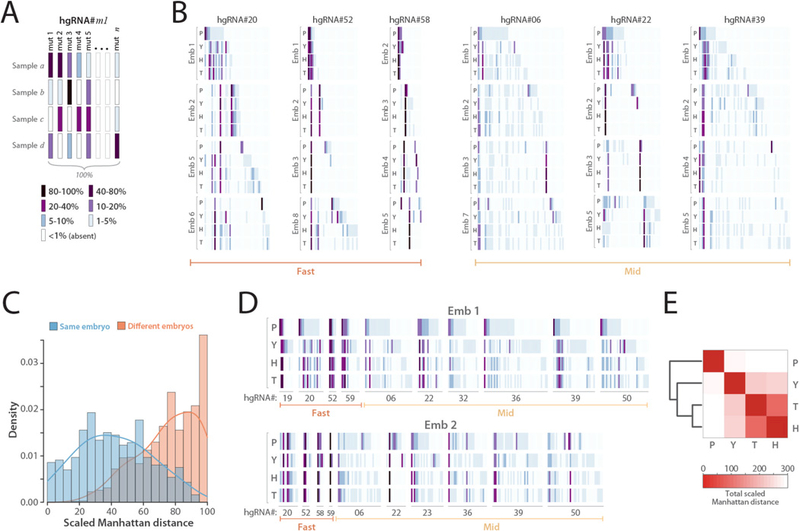
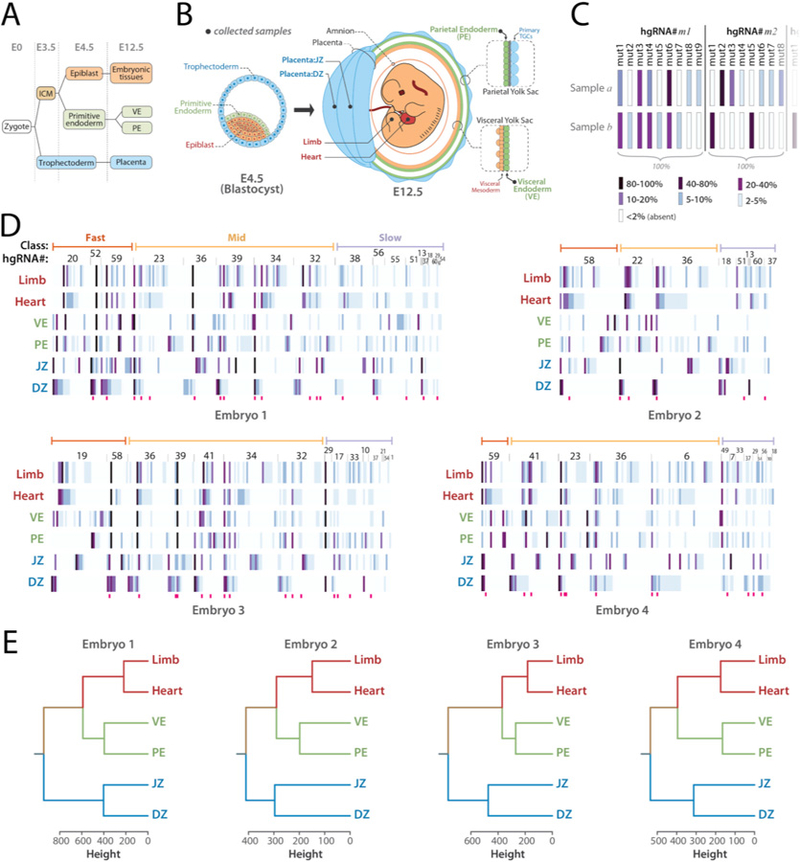
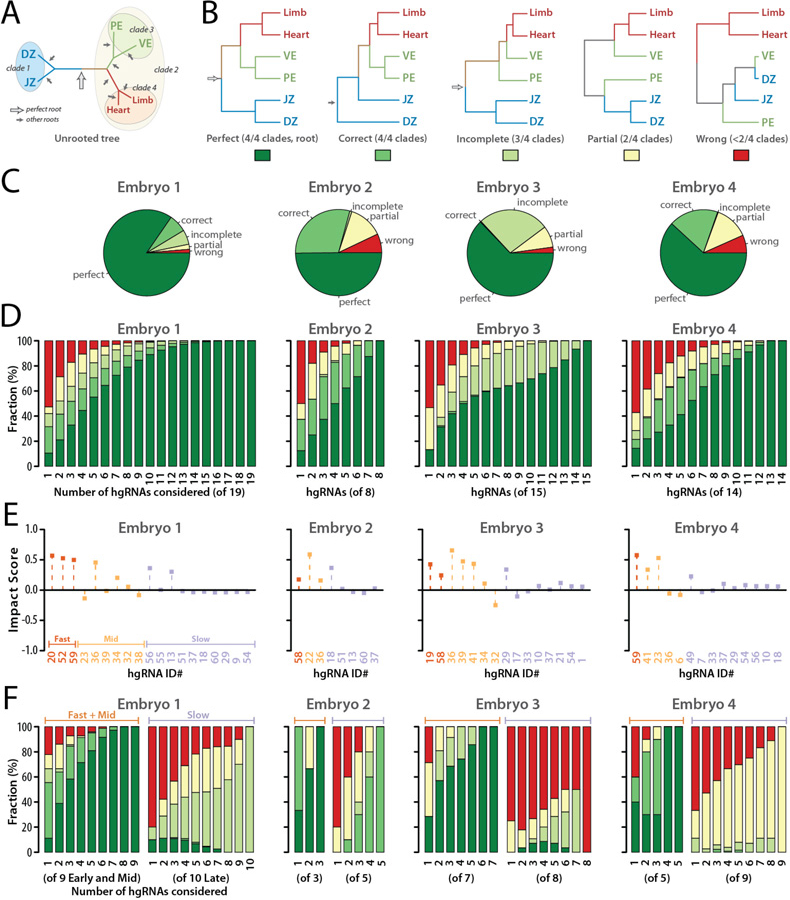
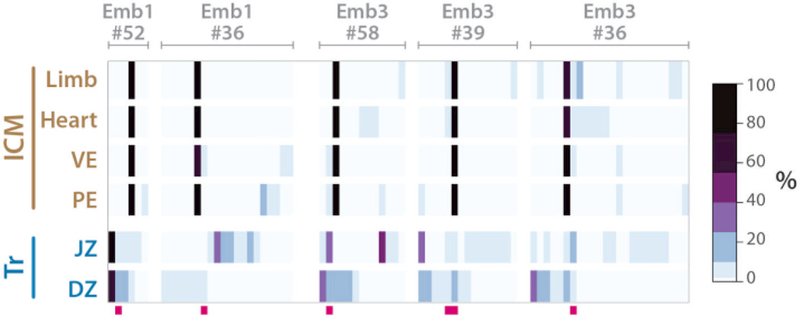
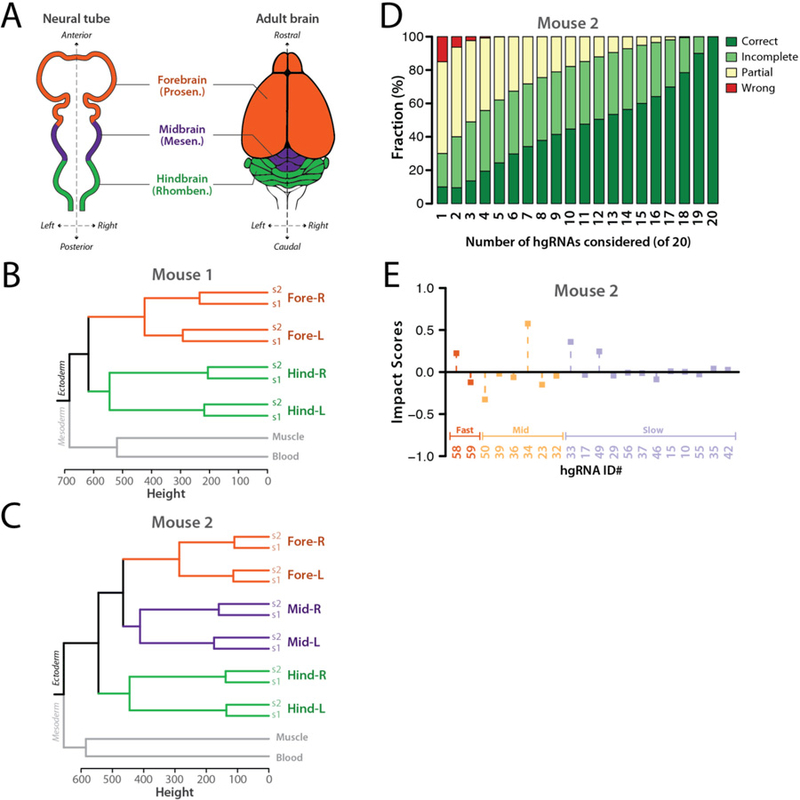
In vivo barcoding using nuclease-induced mutations is a powerful approach for recording biological information, including developmental lineages; however, its application in mammalian systems has been limited. We present in vivo barcoding in the mouse with multiple homing guide RNAs that each generate hundreds of mutant alleles and combine to produce an exponential diversity of barcodes. Activation upon conception and continued mutagenesis through gestation resulted in developmentally barcoded mice wherein information is recorded in lineage-specific mutations. We used these recordings for reliable post hoc reconstruction of the earliest lineages and investigation of axis development in the brain. Our results provide an enabling and versatile platform for in vivo barcoding and lineage tracing in a mammalian model system.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
Comment in
-
In vivo lineage tracing in mice.Nat Rev Genet. 2018 Nov;19(11):669. doi: 10.1038/s41576-018-0048-4. Nat Rev Genet. 2018. PMID: 30135477 No abstract available.
-
The continuously evolving CRISPR barcoding toolbox.Genome Biol. 2018 Sep 25;19(1):143. doi: 10.1186/s13059-018-1541-y. Genome Biol. 2018. PMID: 30253789 Free PMC article.
References
-
- Kaufman MH, The Atlas of Mouse Development (1992).
-
- Sulston JE, Schierenberg E, White JG, Thomson JN, The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol 100, 64–119 (1983). - PubMed
-
- Riddle DL, Blumenthal T, Meyer BJ, Priess JR, in C. elegans II 2nd edition (Cold Spring Harbor Laboratory Press, 1997). - PubMed
-
- Weisblat DA, Sawyer RT, Stent GS, Cell lineage analysis by intracellular injection of a tracer enzyme. Science 202, 1295–1298 (1978). - PubMed
-
- Walsh C, Cepko CL, Widespread dispersion of neuronal clones across functional regions of the cerebral cortex. Science 255, 434–440 (1992). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
