

Interleukin (IL)-2 Is a Key Regulator of T Helper 1 and T Helper 2 Cytokine Expression in Fish: Functional Characterization of Two Divergent IL2 Paralogs in Salmonids
- PMID: 30093902
- PMCID: PMC6070626
- DOI: 10.3389/fimmu.2018.01683
Interleukin (IL)-2 Is a Key Regulator of T Helper 1 and T Helper 2 Cytokine Expression in Fish: Functional Characterization of Two Divergent IL2 Paralogs in Salmonids
Abstract
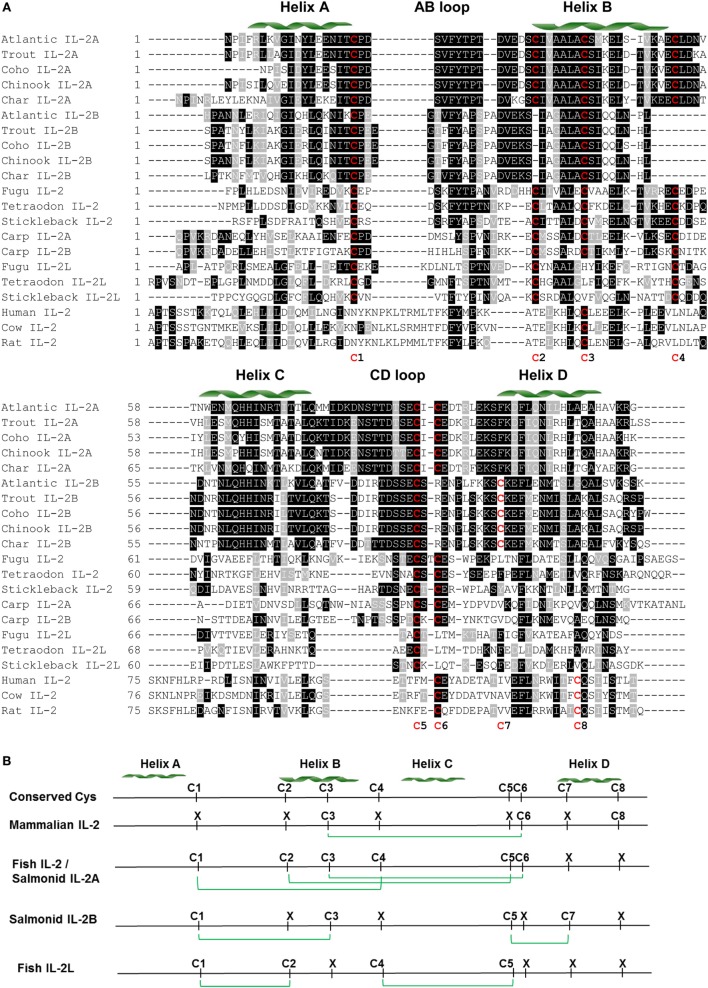
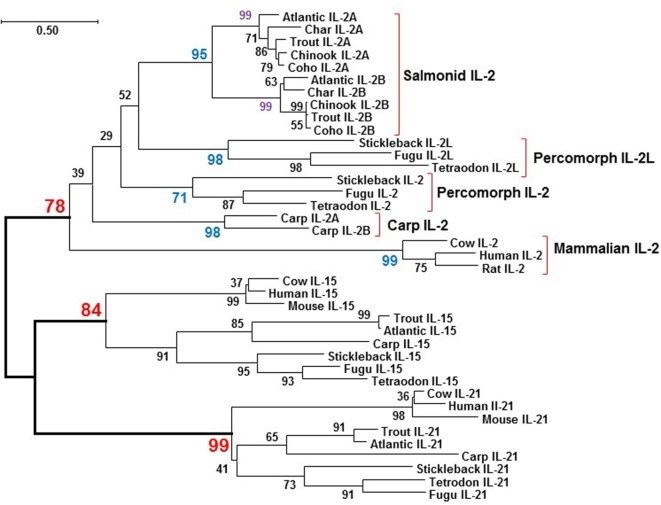
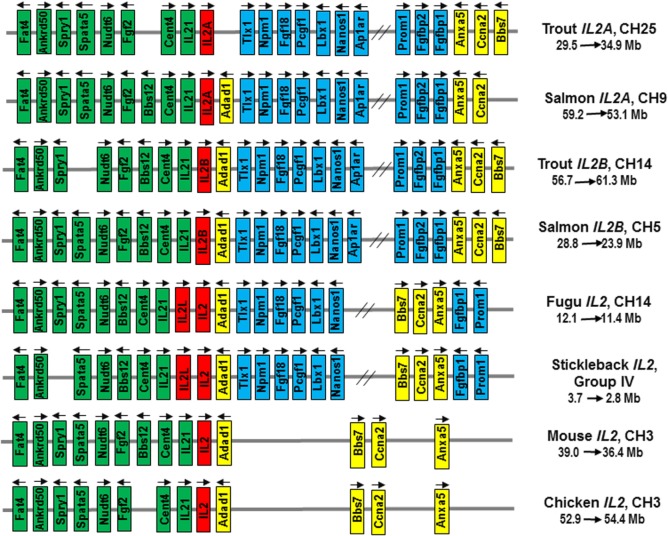
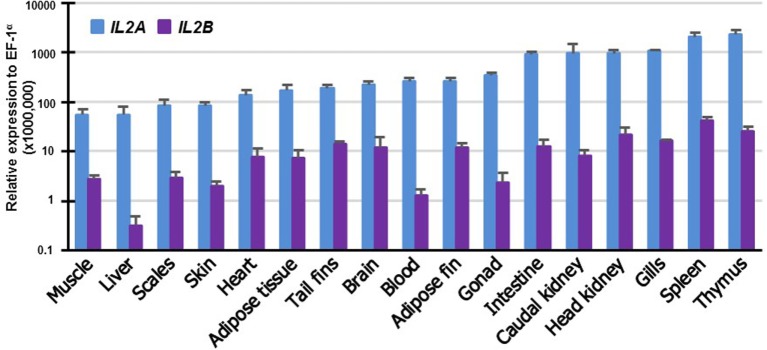
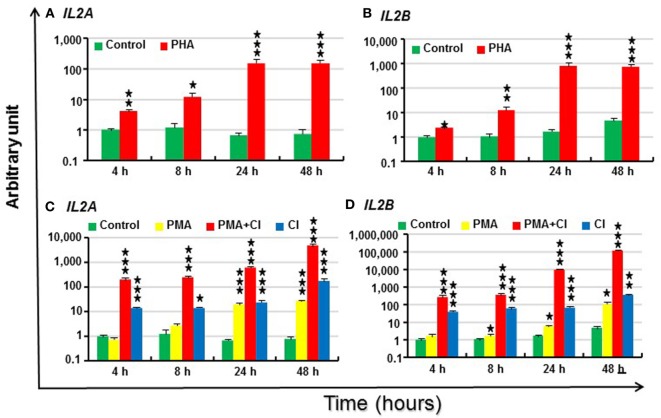
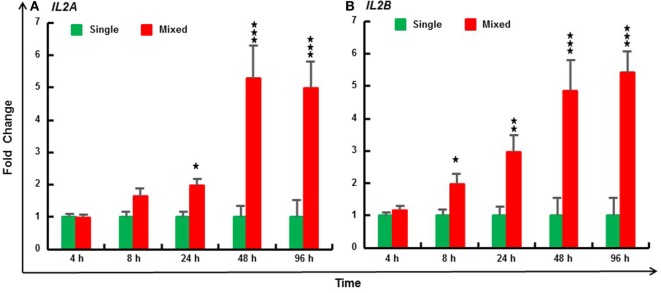
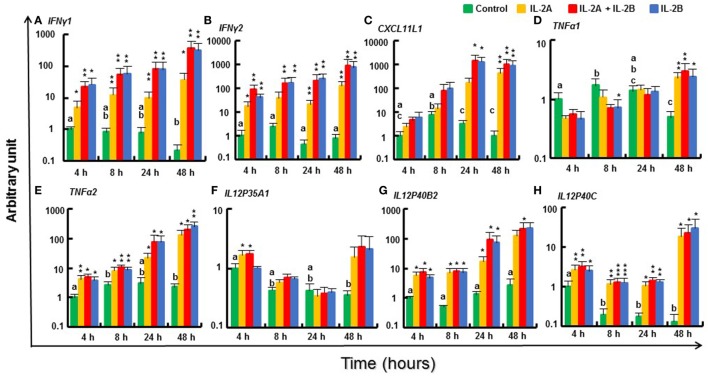
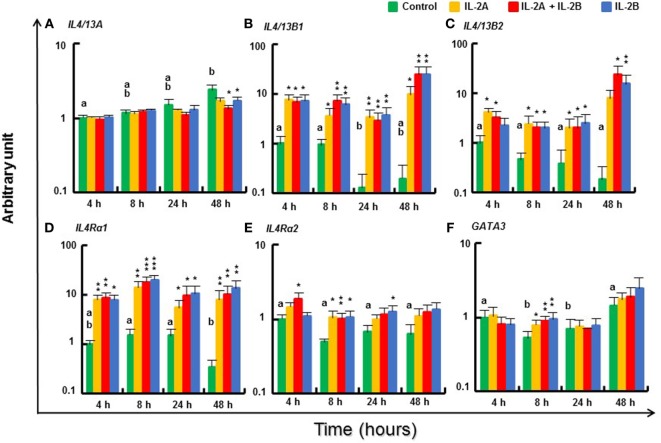
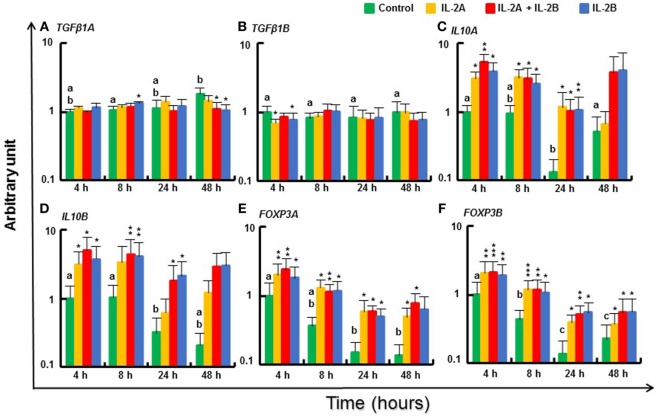
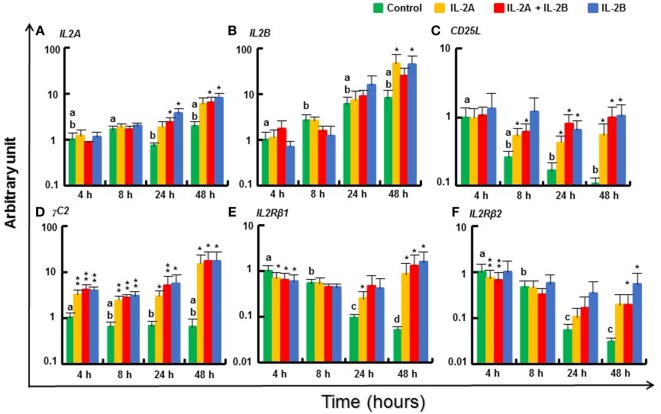
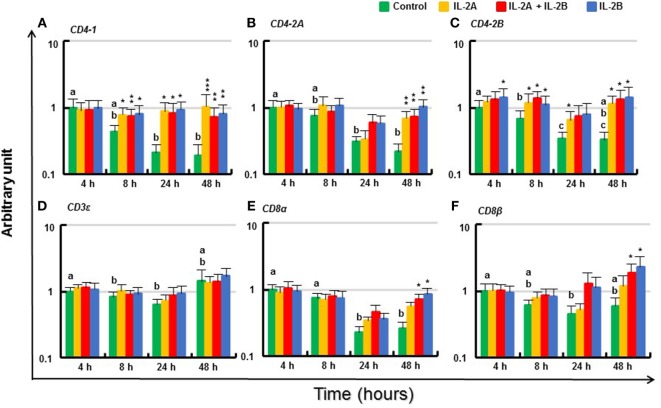
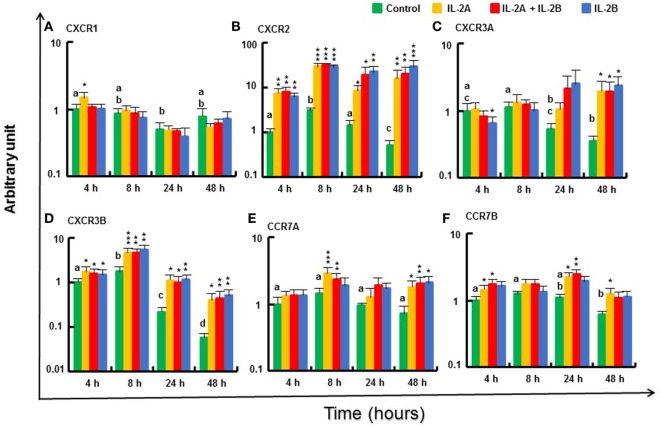
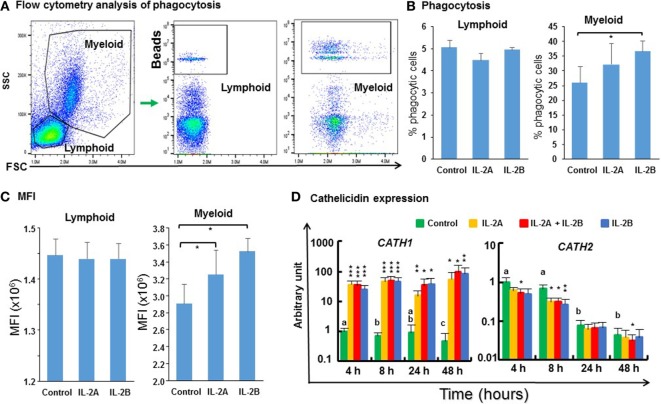
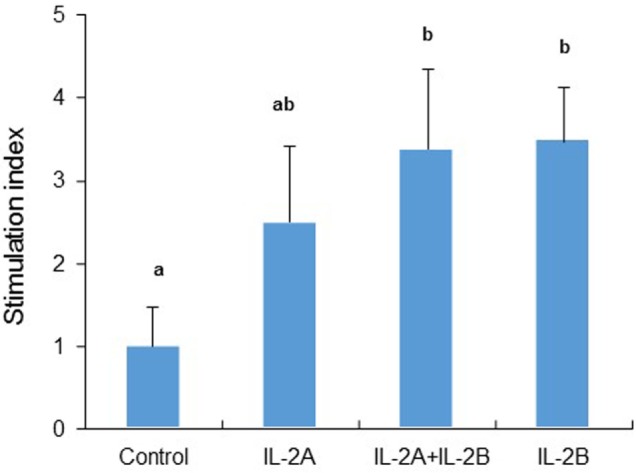
Mammalian interleukin (IL)-2 is a cytokine centrally involved in the differentiation and survival of CD4+ T helper subsets and CD4+ T regulatory cells and in activation of cytotoxic effector lymphocytes. In bony fish, IL2 orthologs have been identified with an additional divergent IL2-Like gene on the same locus present in several fish species. We report here two divergent IL2 paralogs, IL2A and IL2B, in salmonids that originated from the whole genome duplication event in this fish lineage. The salmonid IL2 paralogs differ not only in sequence but also in exon sizes. The IL-2 isoforms that are encoded have disparate pI values and may have evolved to preferentially bind specific IL-2 receptors. Rainbow trout IL2 paralogs are highly expressed in thymus, spleen, gills, kidney and intestine, important tissues/organs in fish T cell development and function. Their expression in peripheral blood leukocytes (PBL) is low constitutively but can be upregulated by the mixed leukocyte reaction, by the T cell mitogen phytohemagglutinin and by signal mimics of T cell activation (phorbol 12-myristate 13-acetate and calcium ionophore). Both trout IL-2 isoforms promoted PBL proliferation and sustained high-level expression of CD4 and CD8, suggesting that trout IL-2 isoforms are T cell growth/survival factors mainly expressed by activated T cells. The recombinant proteins for these two trout IL2 paralogs have been produced in E. coli and possess shared but also distinct bioactivities. IL-2A, but not IL-2B, induced IL12P35A1 and CXCR1 expression in PBL. IL-2B had a stronger effect on upregulation of the T helper 1 (Th1) cytokine interferon-γ (IFNγ) and could sustain CD8α and CD8β expression levels. Nevertheless, both cytokines upregulated key Th1 (IFNγ1, IFNγ2, TNFα2 and IL12) and T helper 2 (Th2) cytokines (IL4/13B1 and IL4/13B2), cytokine and chemokine receptors and the antimicrobial peptide cathelicidin-1 but had limited effects on T helper 17 cytokines and TGFβ1 in PBL. They could also enhance PBL phagocytosis. These results suggest, for the first time in fish, that IL-2 isoforms may have an important role in regulating Th1 and Th2 cell development, and innate and adaptive host defenses in fish, and shed light on lineage-specific expansion, evolution, and functional diversification of IL2 in vertebrates.
Keywords: T cell growth factor; T helper 1; T helper 2; bioactivity; expression; interleukin-2; phagocytosis; salmonids.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous