

Rhodobacterales use a unique L-threonine kinase for the assembly of the nucleotide loop of coenzyme B12
- PMID: 30098062
- PMCID: PMC6195846
- DOI: 10.1111/mmi.14100
Rhodobacterales use a unique L-threonine kinase for the assembly of the nucleotide loop of coenzyme B12
Abstract
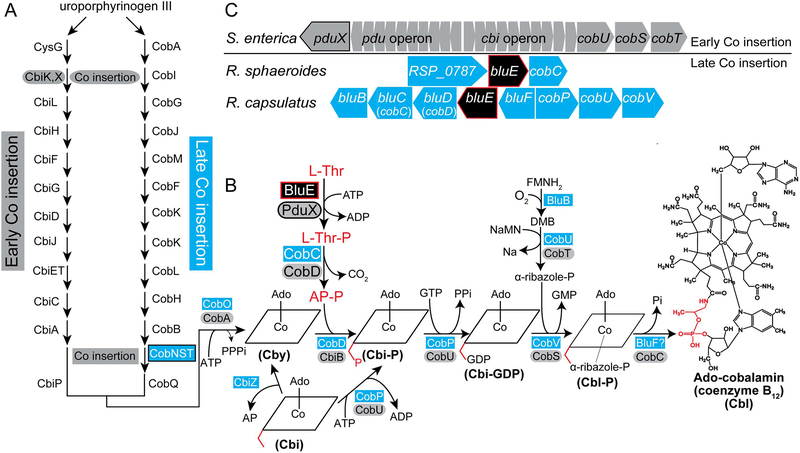
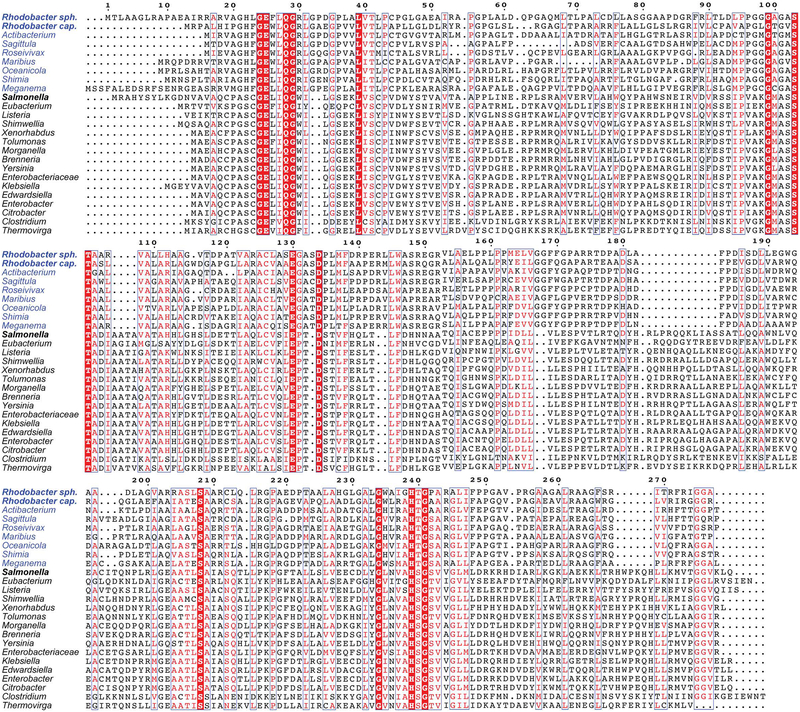
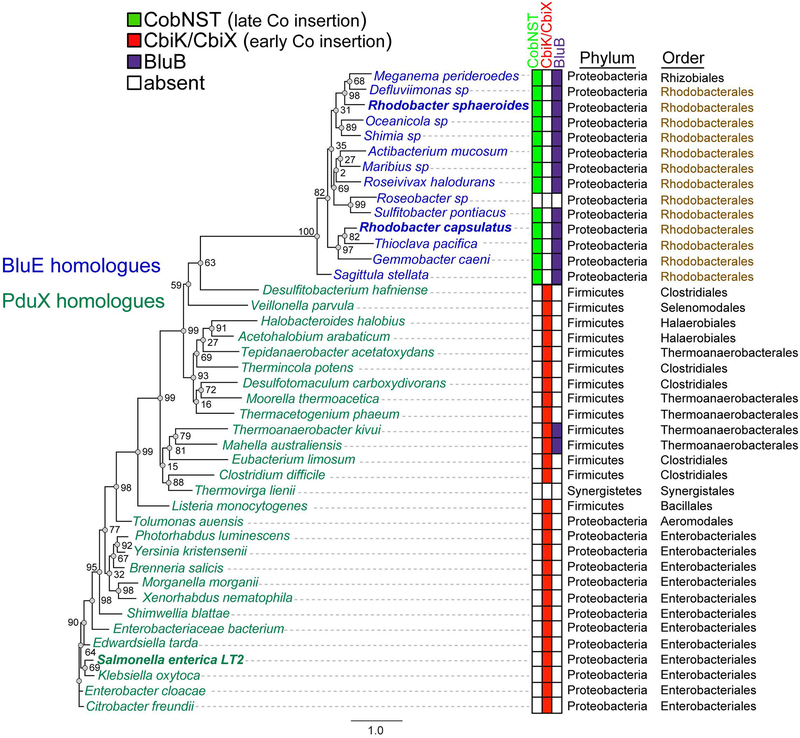
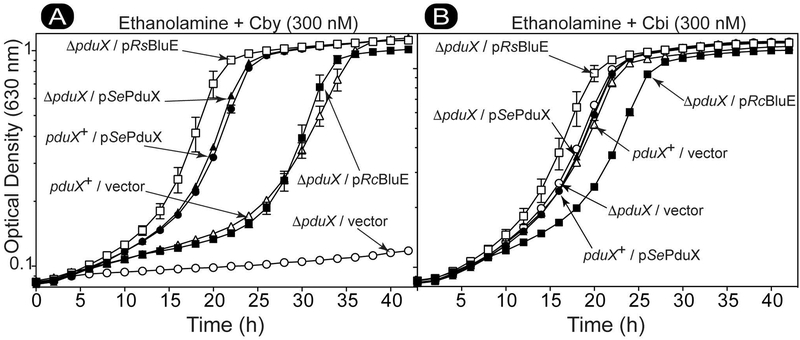
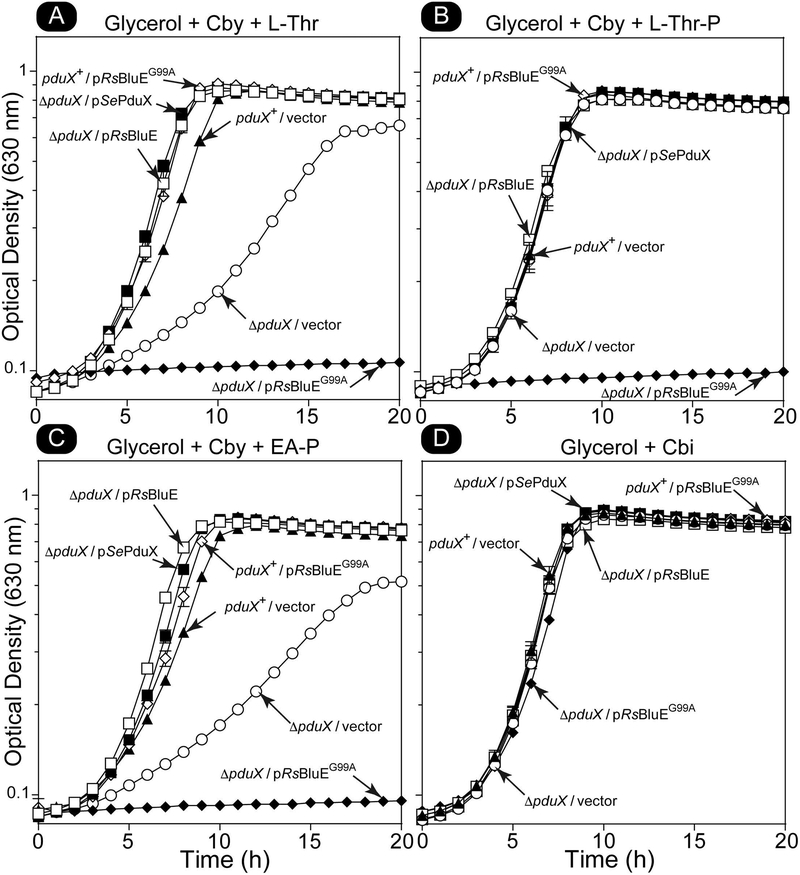
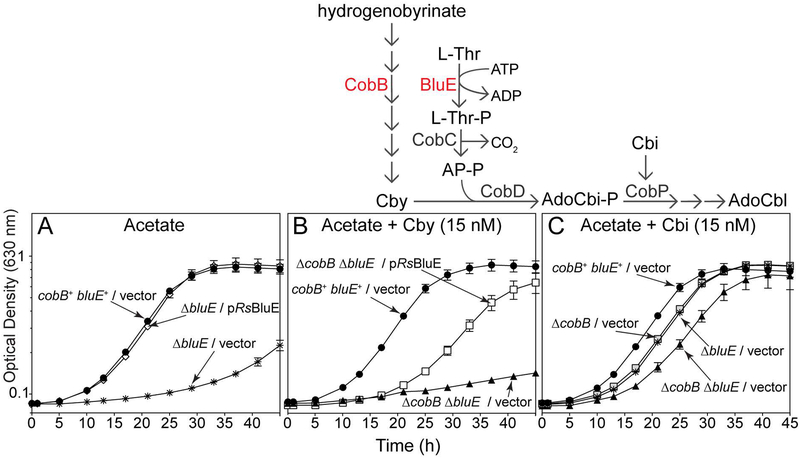
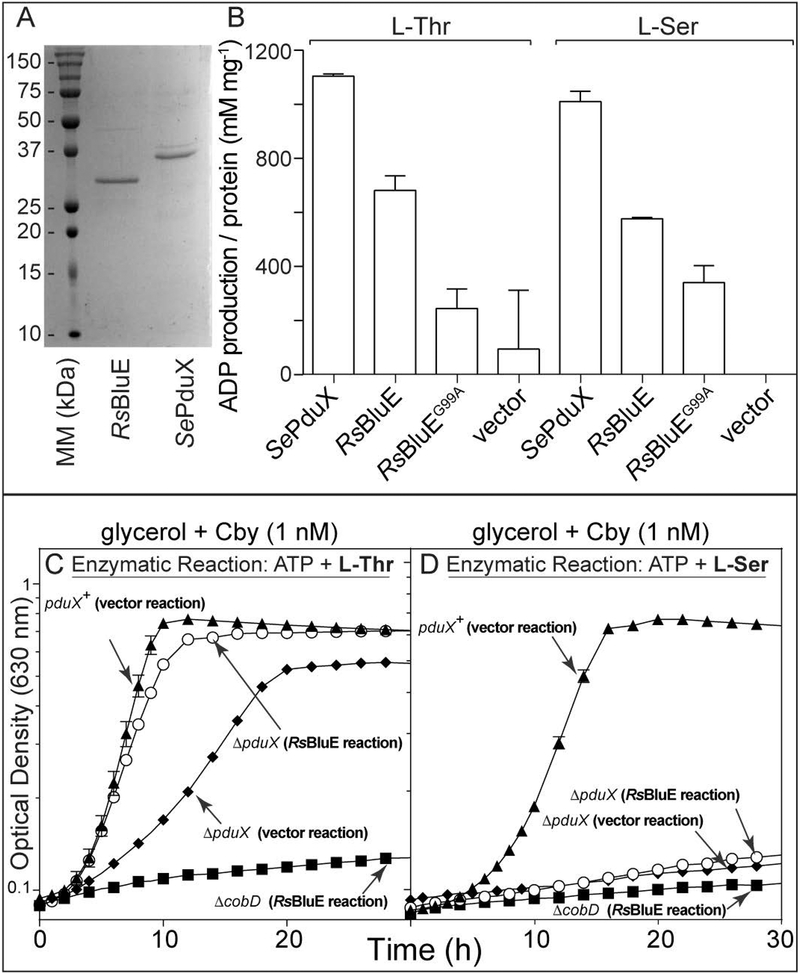
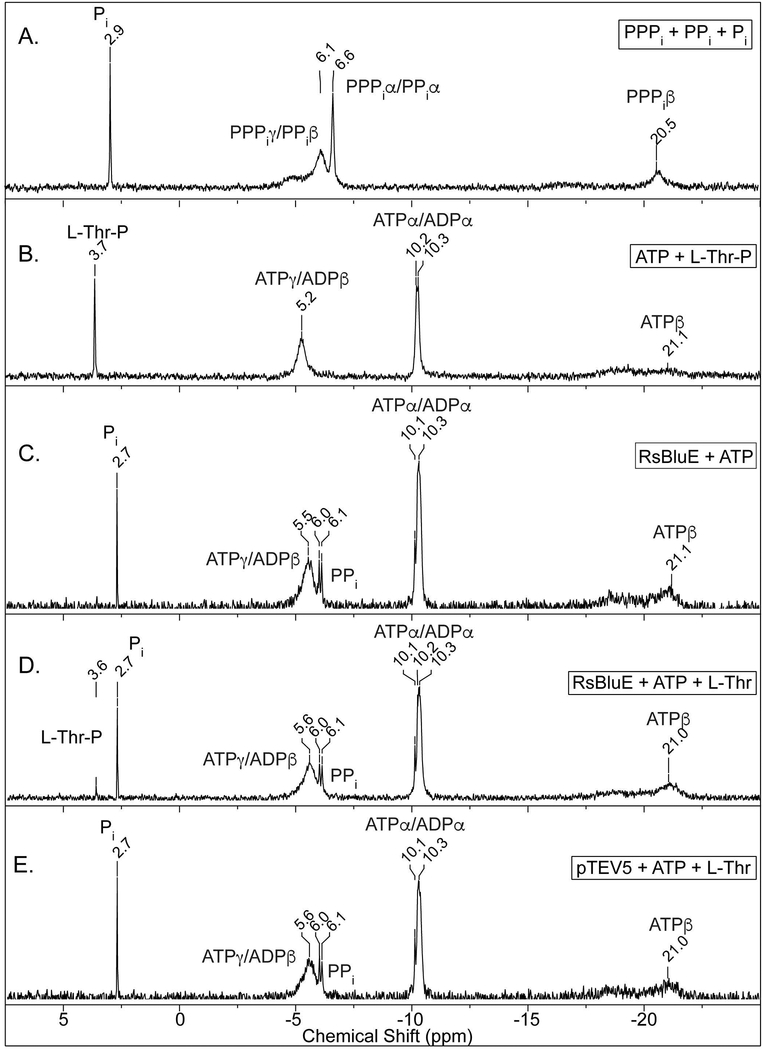
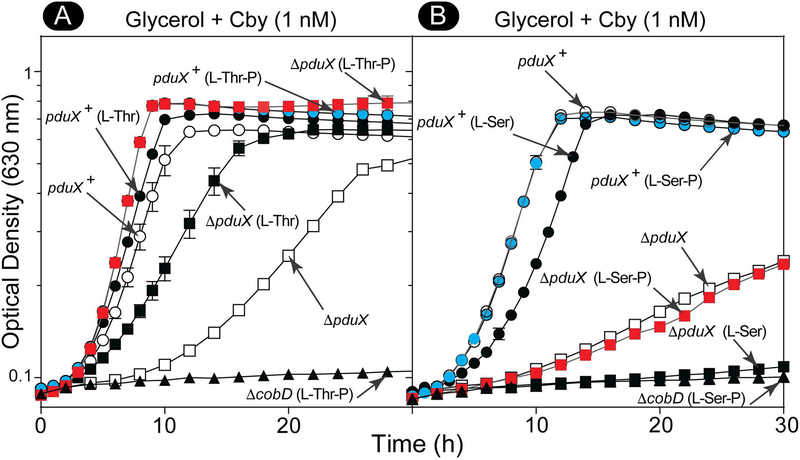
Several of the enzymes involved in the conversion of adenosylcobyric acid (AdoCby) to adenosylcobamide (AdoCba) are yet to be identified and characterized in some cobamide (Cba)-producing prokaryotes. Using a bioinformatics approach, we identified the bluE gene (locus tag RSP_0788) of Rhodobacter sphaeroides 2.4.1 as a putative functional homolog of the L-threonine kinase enzyme (PduX, EC 2.7.1.177) of S. enterica. In AdoCba, (R)-1-aminopropan-2-ol O-phosphate (AP-P) links the nucleotide loop to the corrin ring; most known AdoCba producers derive AP-P from L-Thr-O-3-phosphate (L-Thr-P). Here, we show that RsBluE has L-Thr-independent ATPase activity in vivo and in vitro. We used 31 P-NMR spectroscopy to show that RsBluE generates L-Thr-P at the expense of ATP and is unable to use L-Ser as a substrate. BluE from R. sphaeroides or Rhodobacter capsulatus restored AdoCba biosynthesis in S. enterica ΕpduX and R. sphaeroides ΕbluE mutant strains. R. sphaeroides ΕbluE strains exhibited a decreased pigment phenotype that was restored by complementation with BluE. Finally, phylogenetic analyses revealed that bluE was restricted to the genomes of a few Rhodobacterales that appear to have a preference for a specific form of Cba, namely Coᴽ-(ᴽ-5,6-dimethylbenzimidazolyl-Coᵦ-adenosylcobamide (a.k.a. adenosylcobalamin, AdoCbl; coenzyme B12 , CoB12 ).
© 2018 John Wiley & Sons Ltd.
Figures
Similar articles
-
The PduX enzyme of Salmonella enterica is an L-threonine kinase used for coenzyme B12 synthesis.J Biol Chem. 2008 Apr 25;283(17):11322-9. doi: 10.1074/jbc.M800287200. Epub 2008 Feb 28. J Biol Chem. 2008. PMID: 18308727
-
The SMUL_1544 Gene Product Governs Norcobamide Biosynthesis in the Tetrachloroethene-Respiring Bacterium Sulfurospirillum multivorans.J Bacteriol. 2016 Jul 28;198(16):2236-43. doi: 10.1128/JB.00289-16. Print 2016 Aug 15. J Bacteriol. 2016. PMID: 27274028 Free PMC article.
-
In vivo analysis of cobinamide salvaging in Rhodobacter sphaeroides strain 2.4.1.J Bacteriol. 2009 Jun;191(12):3842-51. doi: 10.1128/JB.00230-09. Epub 2009 Apr 17. J Bacteriol. 2009. PMID: 19376876 Free PMC article.
-
Photosynthetic electron transport and anaerobic metabolism in purple non-sulfur phototrophic bacteria.Antonie Van Leeuwenhoek. 1994;66(1-3):151-64. doi: 10.1007/BF00871637. Antonie Van Leeuwenhoek. 1994. PMID: 7747929 Review.
-
Corrinoid salvaging and cobamide remodeling in bacteria and archaea.J Bacteriol. 2024 Nov 21;206(11):e0028624. doi: 10.1128/jb.00286-24. Epub 2024 Oct 15. J Bacteriol. 2024. PMID: 39404452 Free PMC article. Review.
Cited by
-
Sharing vitamins: Cobamides unveil microbial interactions.Science. 2020 Jul 3;369(6499):eaba0165. doi: 10.1126/science.aba0165. Science. 2020. PMID: 32631870 Free PMC article. Review.
-
Microbial and Genetic Resources for Cobalamin (Vitamin B12) Biosynthesis: From Ecosystems to Industrial Biotechnology.Int J Mol Sci. 2021 Apr 26;22(9):4522. doi: 10.3390/ijms22094522. Int J Mol Sci. 2021. PMID: 33926061 Free PMC article. Review.
-
The dabABC operon is a marker of C4-alkylated monobactam biosynthesis and responsible for (2S,3R)-diaminobutyrate production.iScience. 2024 Feb 12;27(3):109202. doi: 10.1016/j.isci.2024.109202. eCollection 2024 Mar 15. iScience. 2024. PMID: 38433893 Free PMC article.
-
Biosynthesis of the modified tetrapyrroles-the pigments of life.J Biol Chem. 2020 May 15;295(20):6888-6925. doi: 10.1074/jbc.REV120.006194. Epub 2020 Apr 2. J Biol Chem. 2020. PMID: 32241908 Free PMC article. Review.
-
The l-Thr Kinase/l-Thr-Phosphate Decarboxylase (CobD) Enzyme from Methanosarcina mazei Gö1 Contains Metallocenters Needed for Optimal Activity.Biochemistry. 2019 Jul 30;58(30):3260-3279. doi: 10.1021/acs.biochem.9b00268. Epub 2019 Jul 17. Biochemistry. 2019. PMID: 31268299 Free PMC article.
References
-
- Amin SA, Green DH, Gardes A, Romano A, Trimble L, and Carrano CJ (2012) Siderophore-mediated iron uptake in two clades of Marinobacter spp. associated with phytoplankton: the role of light. Biometals 25: 181–192. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
