

Cell cycle checkpoint control: The cyclin G1/Mdm2/p53 axis emerges as a strategic target for broad-spectrum cancer gene therapy - A review of molecular mechanisms for oncologists
- PMID: 30101008
- PMCID: PMC6083405
- DOI: 10.3892/mco.2018.1657
Cell cycle checkpoint control: The cyclin G1/Mdm2/p53 axis emerges as a strategic target for broad-spectrum cancer gene therapy - A review of molecular mechanisms for oncologists
Abstract
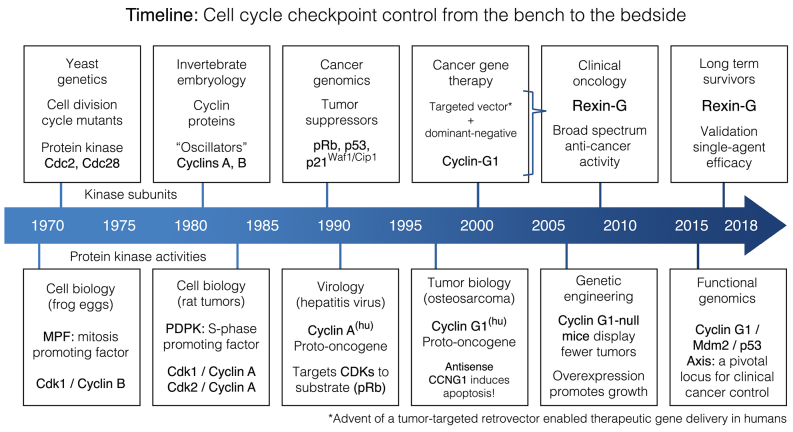
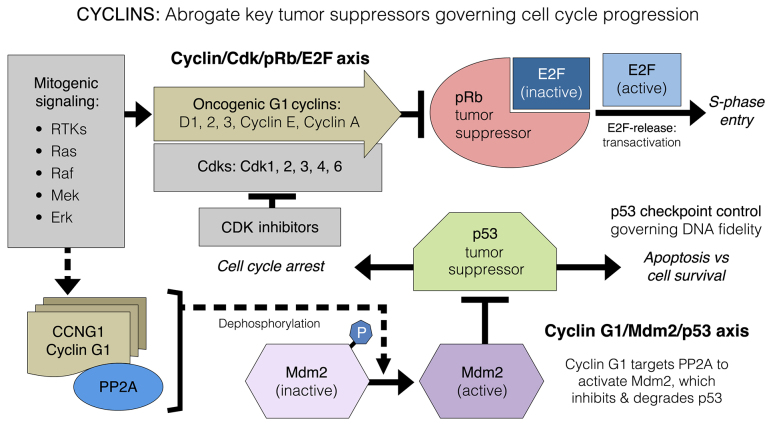
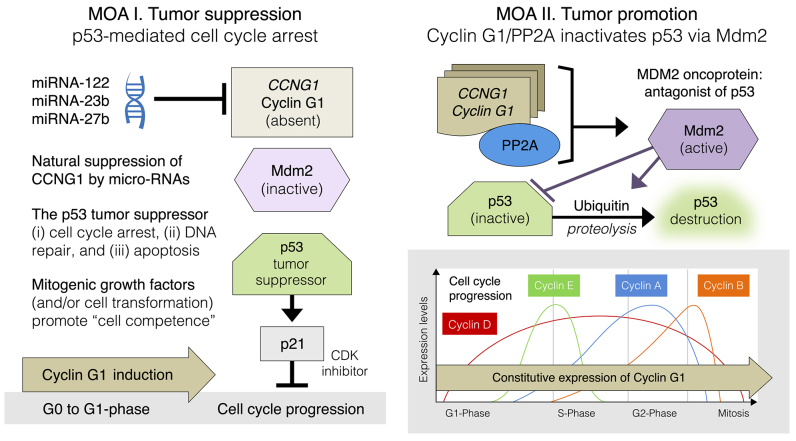
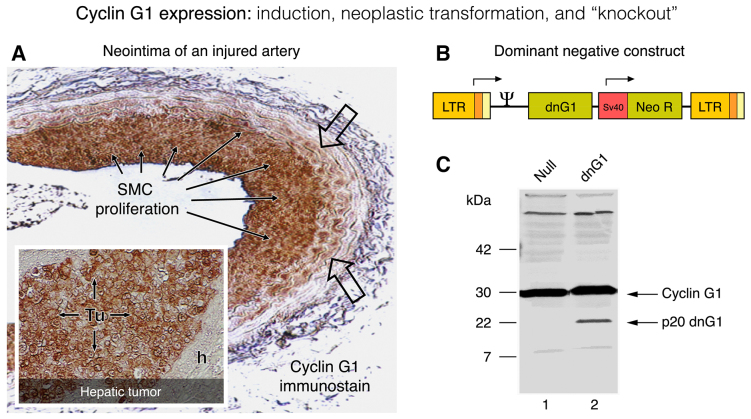
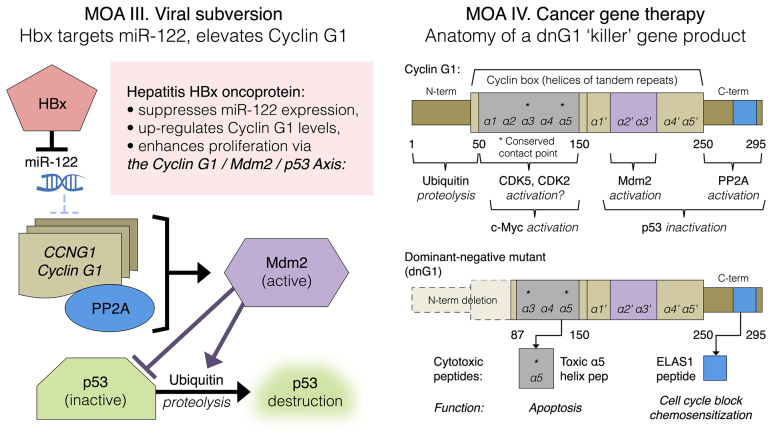
Basic research in genetics, biochemistry and cell biology has identified the executive enzymes and protein kinase activities that regulate the cell division cycle of all eukaryotic organisms, thereby elucidating the importance of site-specific protein phosphorylation events that govern cell cycle progression. Research in cancer genomics and virology has provided meaningful links to mammalian checkpoint control elements with the characterization of growth-promoting proto-oncogenes encoding c-Myc, Mdm2, cyclins A, D1 and G1, and opposing tumor suppressor proteins, such as p53, pRb, p16INK4A and p21WAF1, which are commonly dysregulated in cancer. While progress has been made in identifying numerous enzymes and molecular interactions associated with cell cycle checkpoint control, the marked complexity, particularly the functional redundancy, of these cell cycle control enzymes in mammalian systems, presents a major challenge in discerning an optimal locus for therapeutic intervention in the clinical management of cancer. Recent advances in genetic engineering, functional genomics and clinical oncology converged in identifying cyclin G1 (CCNG1 gene) as a pivotal component of a commanding cyclin G1/Mdm2/p53 axis and a strategic locus for re-establishing cell cycle control by means of therapeutic gene transfer. The purpose of the present study is to provide a focused review of cycle checkpoint control as a practicum for clinical oncologists with an interest in applied molecular medicine. The aim is to present a unifying model that: i) clarifies the function of cyclin G1 in establishing proliferative competence, overriding p53 checkpoints and advancing cell cycle progression; ii) is supported by studies of inhibitory microRNAs linking CCNG1 expression to the mechanisms of carcinogenesis and viral subversion; and iii) provides a mechanistic basis for understanding the broad-spectrum anticancer activity and single-agent efficacy observed with dominant-negative cyclin G1, whose cytocidal mechanism of action triggers programmed cell death. Clinically, the utility of companion diagnostics for cyclin G1 pathways is anticipated in the staging, prognosis and treatment of cancers, including the potential for rational combinatorial therapies.
Keywords: CCNG1; Mdm2; Rexin-G; cancer gene therapy; cyclin-dependent kinase; gene delivery; microRNAs; p53; protein phosphatase 2A; proto-oncogene; tumor suppressor.
Figures
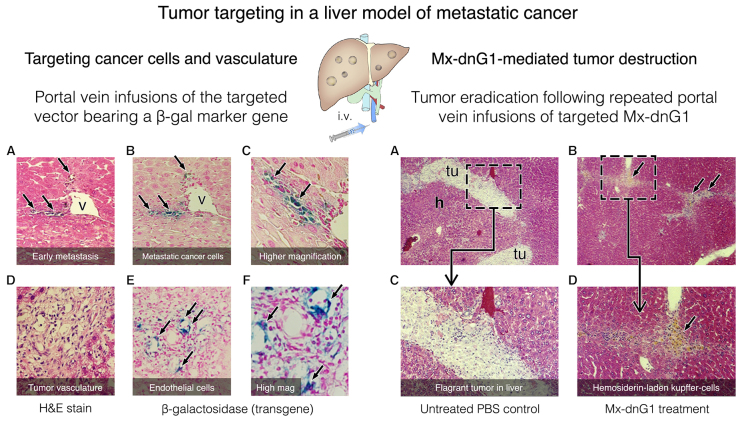
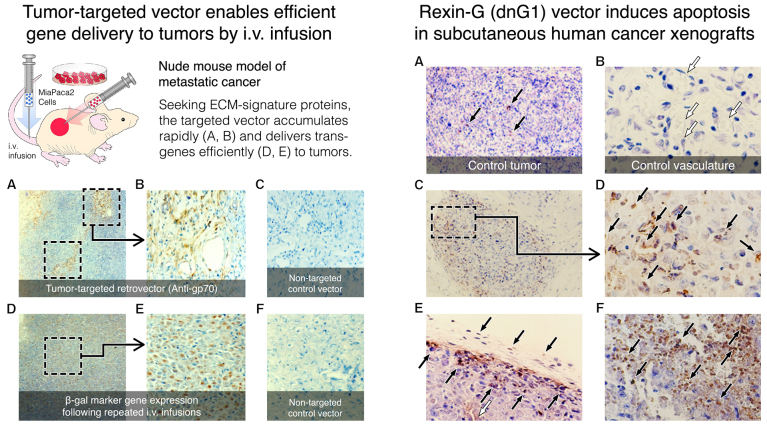
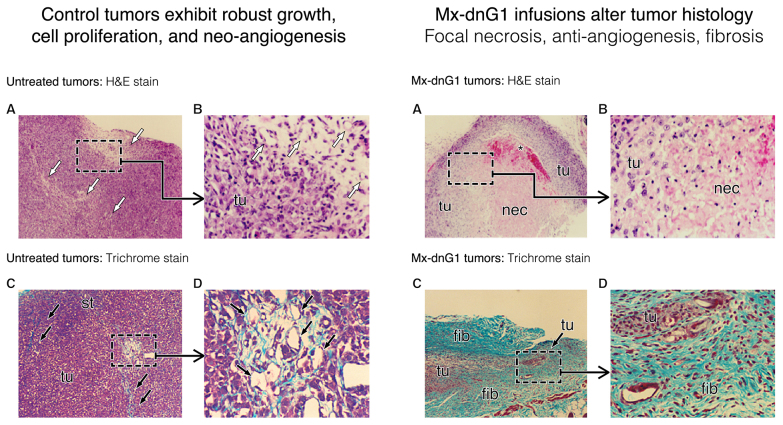
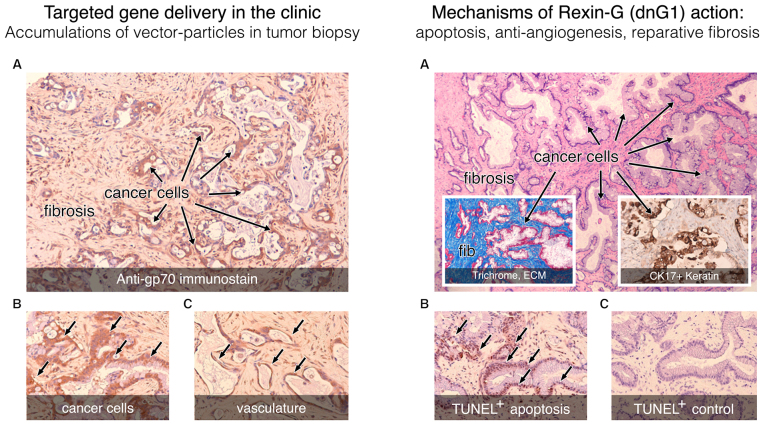
References
-
- Weinstein IB. The origins of human cancer: Molecular mechanisms of carcinogenesis and their implications for cancer prevention and treatment-twenty-seventh G.H.A. Clowes memorial award lecture. Cancer Res. 1988;48:4135–4143. - PubMed
-
- Kikkawa U, Takai Y, Tanaka Y, Miyake R, Nishizuka Y. Protein kinase C as a possible receptor protein of tumor-promoting phorbol esters. J Biol Chem. 1983;258:11442–11445. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous