

Concentrations of oxidized linoleic acid derived lipid mediators in the amygdala and periaqueductal grey are reduced in a mouse model of chronic inflammatory pain
- PMID: 30103924
- PMCID: PMC6269101
- DOI: 10.1016/j.plefa.2018.07.015
Concentrations of oxidized linoleic acid derived lipid mediators in the amygdala and periaqueductal grey are reduced in a mouse model of chronic inflammatory pain
Abstract
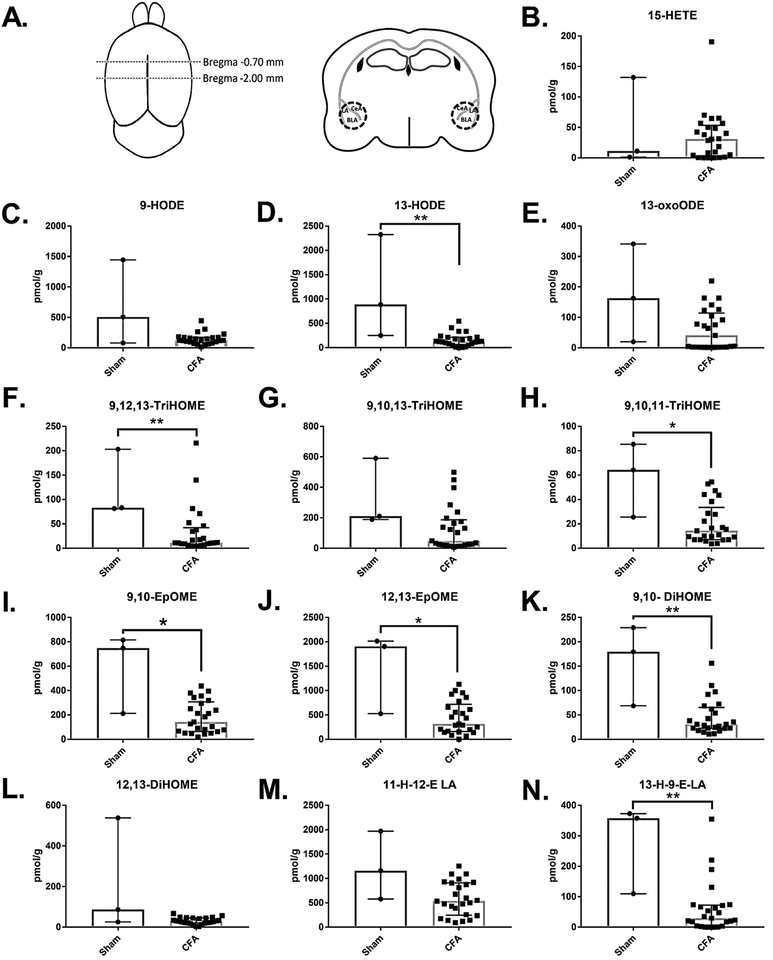
Chronic pain is both a global public health concern and a serious source of personal suffering for which current treatments have limited efficacy. Recently, oxylipins derived from linoleic acid (LA), the most abundantly consumed polyunsaturated fatty acid in the modern diet, have been implicated as mediators of pain in the periphery and spinal cord. However, oxidized linoleic acid derived mediators (OXLAMs) remain understudied in the brain, particularly during pain states. In this study, we employed a mouse model of chronic inflammatory pain followed by a targeted lipidomic analysis of the animals' amygdala and periaqueductal grey (PAG) using LC-MS/MS to investigate the effect of chronic inflammatory pain on oxylipin concentrations in these two brain nuclei known to participate in pain sensation and perception. From punch biopsies of these brain nuclei, we detected twelve OXLAMs in both the PAG and amygdala and one arachidonic acid derived mediator, 15-HETE, in the amygdala only. In the amygdala, we observed an overall decrease in the concentration of the majority of OXLAMs detected, while in the PAG the concentrations of only the epoxide LA derived mediators, 9,10-EpOME and 12,13-EpOME, and one trihydroxy LA derived mediator, 9,10,11-TriHOME, were reduced. This data provides the first evidence that OXLAM concentrations in the brain are affected by chronic pain, suggesting that OXLAMs may be relevant to pain signaling and adaptation to chronic pain in pain circuits in the brain and that the current view of OXLAMs in nociception derived from studies in the periphery is incomplete.
Published by Elsevier Ltd.
Figures


References
-
- Bazinet RP, Layé S, Polyunsaturated fatty acids and their metabolites in brain function and disease, Nat. Rev. Neurosci 15 (2014) 771–785, https://doi.org/10.1038/nrn3820. - DOI - PubMed
-
- Chen CT, Green JT, Orr SK, Bazinet RP, Regulation of brain polyunsaturated fatty acid uptake and turnover, Prostaglandins Leukot Essent Fatty Acids 79 (2008) 85–91, https://doi.org/10.1016/j.plefa.2008.09.003. - DOI - PubMed
-
- Farooqui AA, Horrocks LA, Phospholipase A2-generated lipid mediators in the brain: the good, the bad, and the ugly, Neuroscientist 12 (2006) 245–260, https://doi.org/10.1177/1073858405285923. - DOI - PubMed
-
- Chang C-Y, Ke D-S, Chen J-Y, Essential fatty acids and human brain, Acta Neurol. Taiwan 18 (2009) 231–241. - PubMed
-
- Haag M, Essential fatty acids and the brain, Can. J. Psychiatry 48 (2003) 195–203, https://doi.org/10.1177/070674370304800308. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
