

The Vibrio H-Ring Facilitates the Outer Membrane Penetration of the Polar Sheathed Flagellum
- PMID: 30104237
- PMCID: PMC6182240
- DOI: 10.1128/JB.00387-18
The Vibrio H-Ring Facilitates the Outer Membrane Penetration of the Polar Sheathed Flagellum
Abstract
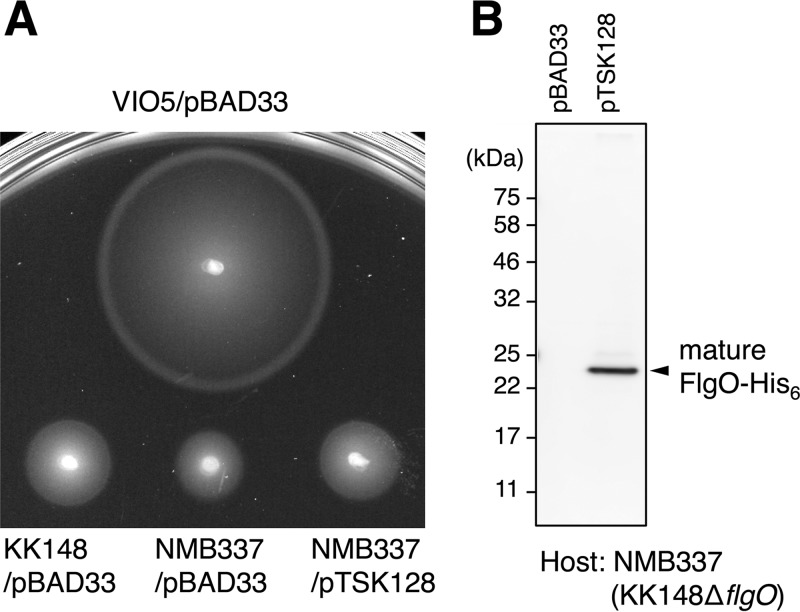
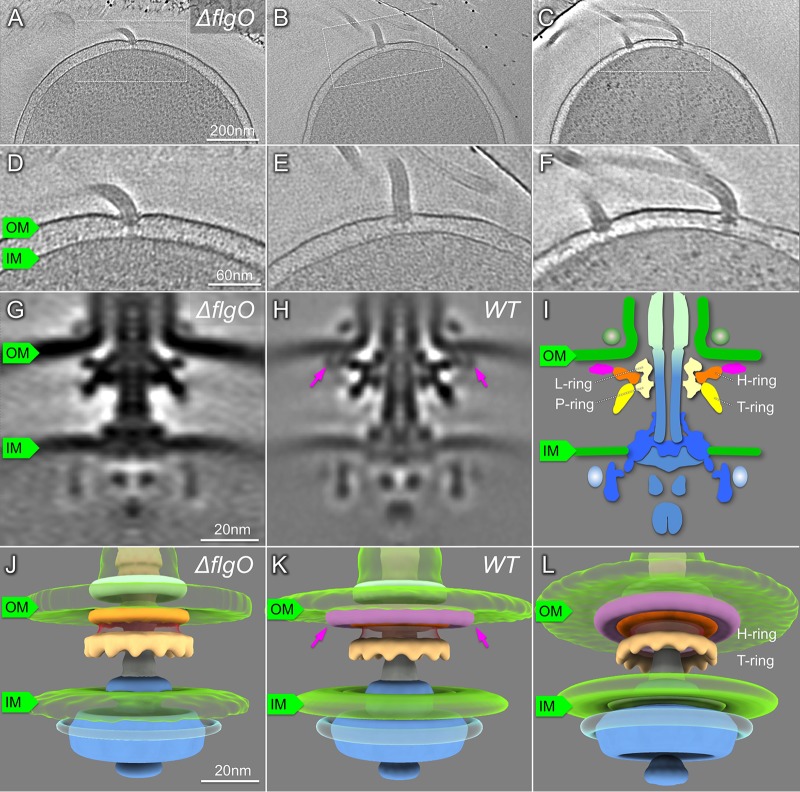
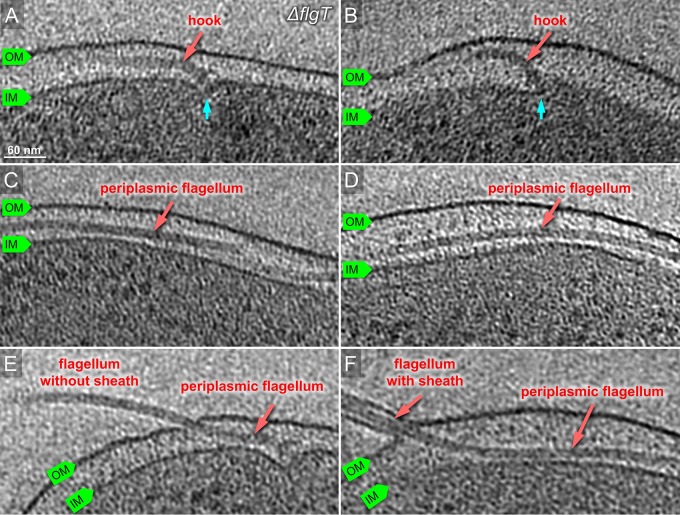
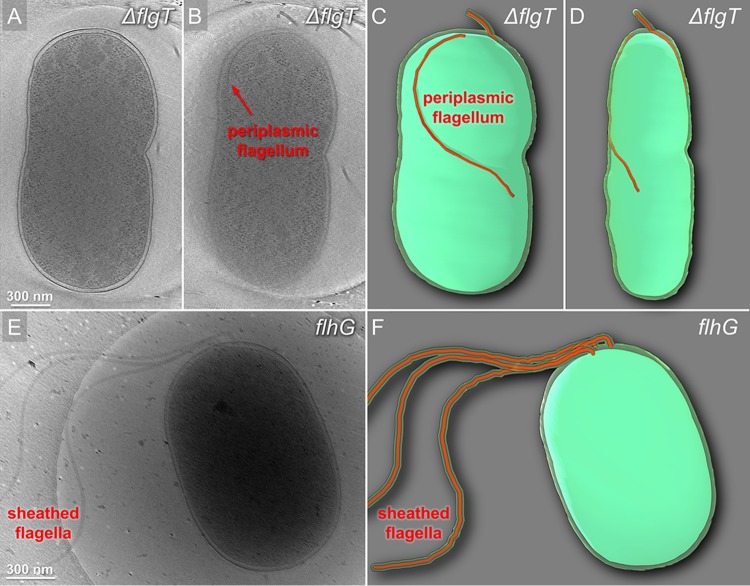
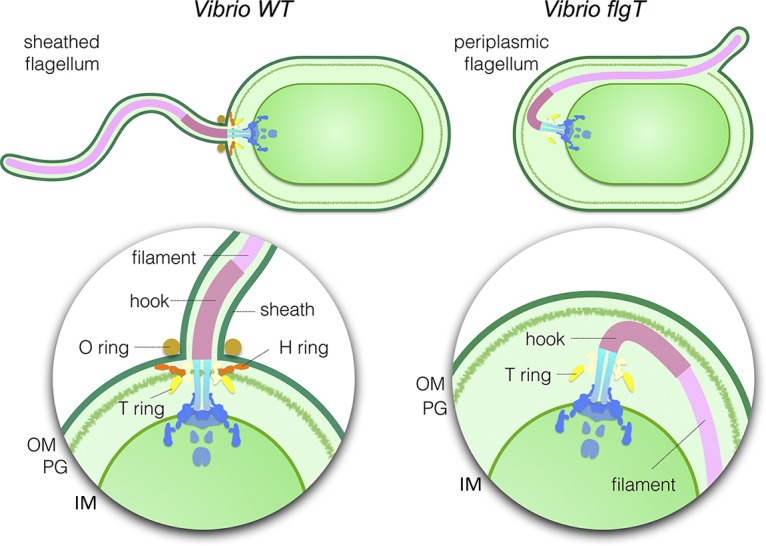
The bacterial flagellum has evolved as one of the most remarkable nanomachines in nature. It provides swimming and swarming motilities that are often essential for the bacterial life cycle and pathogenesis. Many bacteria such as Salmonella and Vibrio species use flagella as an external propeller to move to favorable environments, whereas spirochetes utilize internal periplasmic flagella to drive a serpentine movement of the cell bodies through tissues. Here, we use cryo-electron tomography to visualize the polar sheathed flagellum of Vibrio alginolyticus with particular focus on a Vibrio-specific feature, the H-ring. We characterized the H-ring by identifying its two components FlgT and FlgO. We found that the majority of flagella are located within the periplasmic space in the absence of the H-ring, which are different from those of external flagella in wild-type cells. Our results not only indicate the H-ring has a novel function in facilitating the penetration of the outer membrane and the assembly of the external sheathed flagella but also are consistent with the notion that the flagella have evolved to adapt highly diverse needs by receiving or removing accessary genes.IMPORTANCE Flagellum is the major organelle for motility in many bacterial species. While most bacteria possess external flagella, such as the multiple peritrichous flagella found in Escherichia coli and Salmonella enterica or the single polar sheathed flagellum in Vibrio spp., spirochetes uniquely assemble periplasmic flagella, which are embedded between their inner and outer membranes. Here, we show for the first time that the external flagella in Vibrio alginolyticus can be changed as periplasmic flagella by deleting two flagellar genes. The discovery here may provide new insights into the molecular basis underlying assembly, diversity, and evolution of flagella.
Keywords: flagellar assembly; flagellar evolution; membrane penetration; periplasmic flagella.
Copyright © 2018 American Society for Microbiology.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
