

Bond swapping from a charge cloud allows flexible coordination of upstream signals through WASP: Multiple regulatory roles for the WASP basic region
- PMID: 30104412
- PMCID: PMC6166713
- DOI: 10.1074/jbc.RA118.003290
Bond swapping from a charge cloud allows flexible coordination of upstream signals through WASP: Multiple regulatory roles for the WASP basic region
Abstract
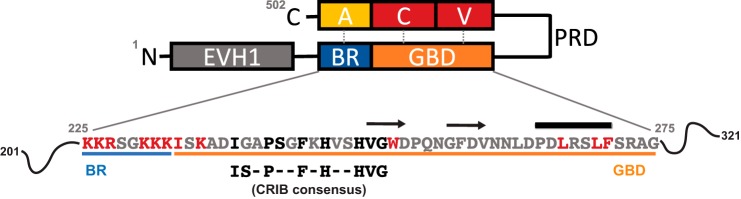
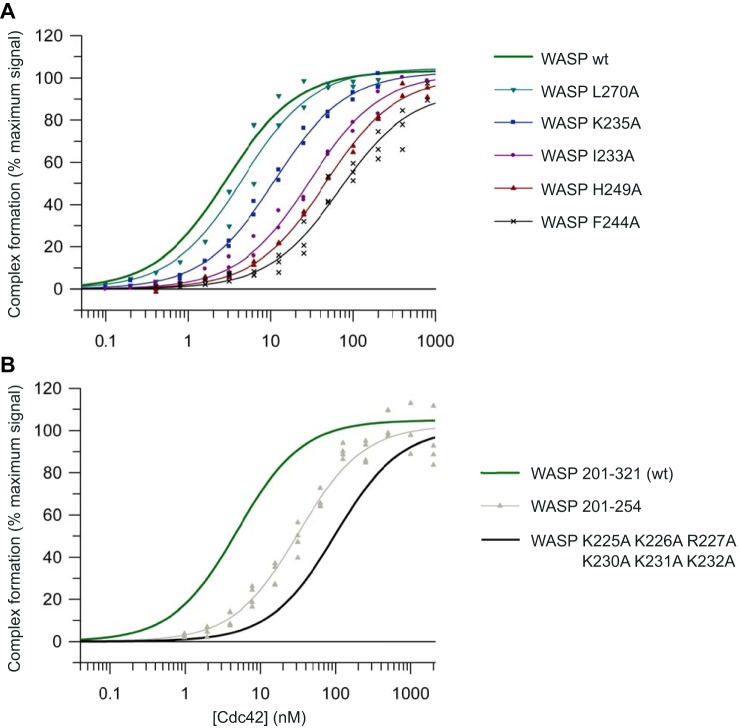
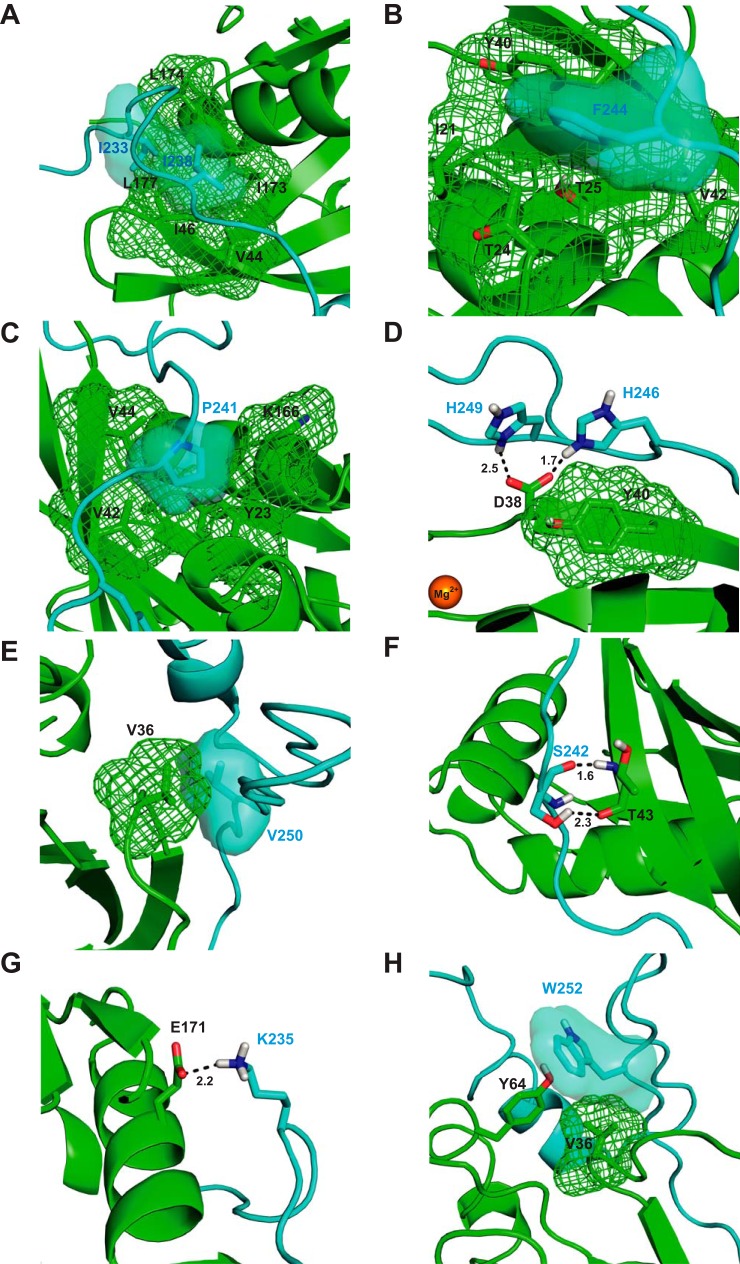
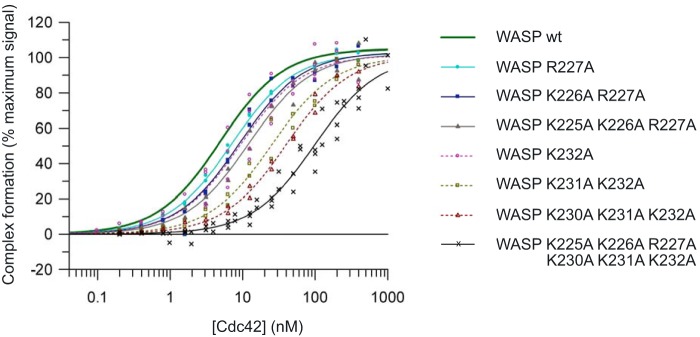
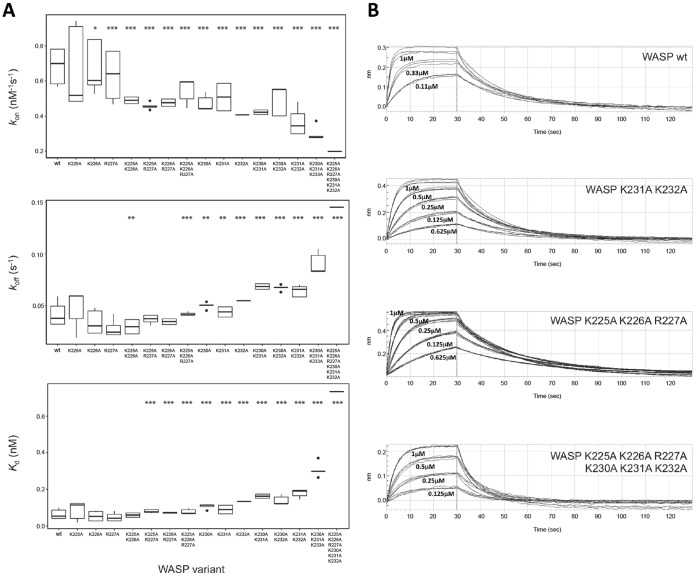
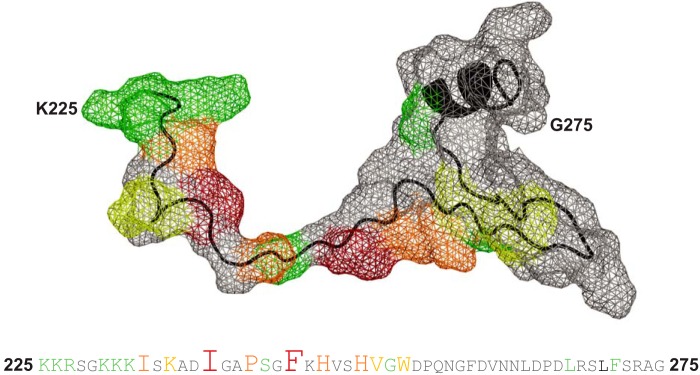
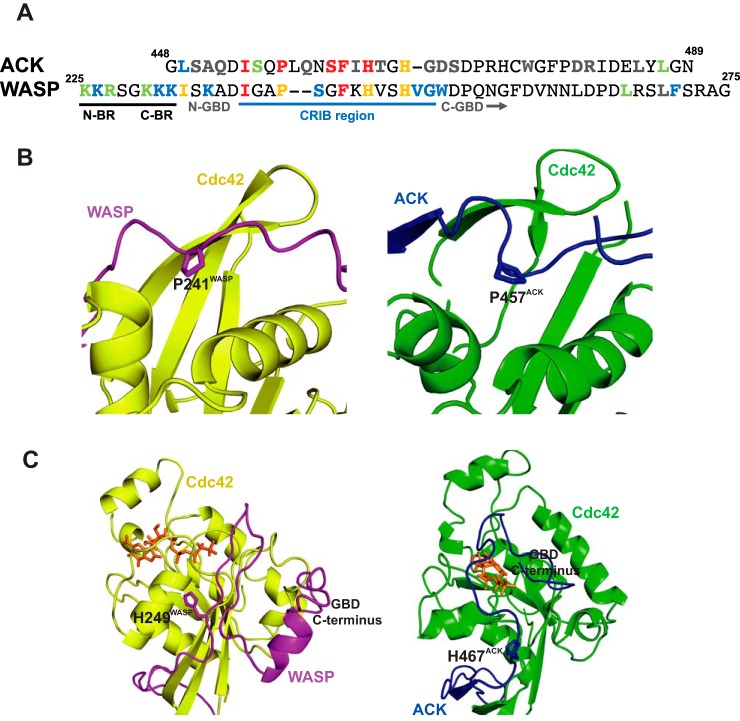
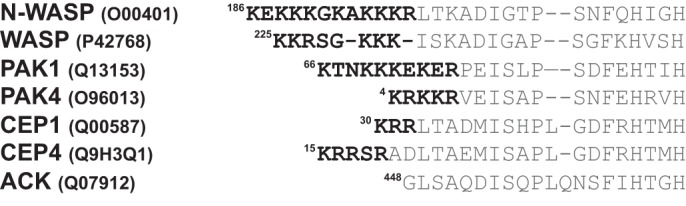
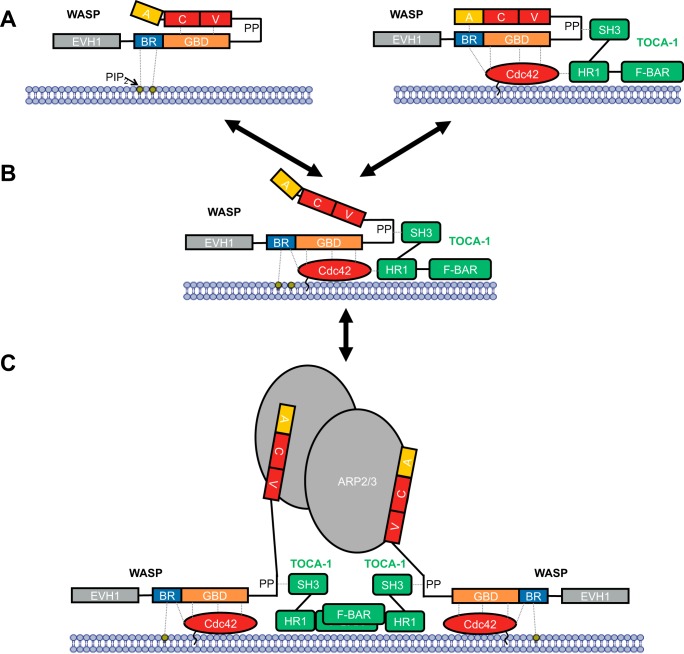
Wiskott-Aldrich syndrome protein (WASP) activates the actin-related protein 2/3 homolog (Arp2/3) complex and regulates actin polymerization in a physiological setting. Cell division cycle 42 (Cdc42) is a key activator of WASP, which binds Cdc42 through a Cdc42/Rac-interactive binding (CRIB)-containing region that defines a subset of Cdc42 effectors. Here, using site-directed mutagenesis and binding affinity determination and kinetic assays, we report the results of an investigation into the energetic contributions of individual WASP residues to both the Cdc42-WASP binding interface and the kinetics of complex formation. Our results support the previously proposed dock-and-coalesce binding mechanism, initiated by electrostatic steering driven by WASP's basic region and followed by a coalescence phase likely driven by the conserved CRIB motif. The WASP basic region, however, appears also to play a role in the final complex, as its mutation affected both on- and off-rates, suggesting a more comprehensive physiological role for this region centered on the C-terminal triad of positive residues. These results highlight the expanding roles of the basic region in WASP and other CRIB-containing effector proteins in regulating complex cellular processes and coordinating multiple input signals. The data presented improve our understanding of the Cdc42-WASP interface and also add to the body of information available for Cdc42-effector complex formation, therapeutic targeting of which has promise for Ras-driven cancers. Our findings suggest that combining high-affinity peptide-binding sequences with short electrostatic steering sequences could increase the efficacy of peptidomimetic candidates designed to interfere with Cdc42 signaling in cancer.
Keywords: CDC42; cell signaling; cytoskeleton; intrinsically disordered protein; protein conformation; protein motif; protein–protein interaction; small GTPase; structure–function; thermodynamics.
© 2018 Tetley et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
