

Spatial Representations in the Human Brain
- PMID: 30104966
- PMCID: PMC6078001
- DOI: 10.3389/fnhum.2018.00297
Spatial Representations in the Human Brain
Abstract
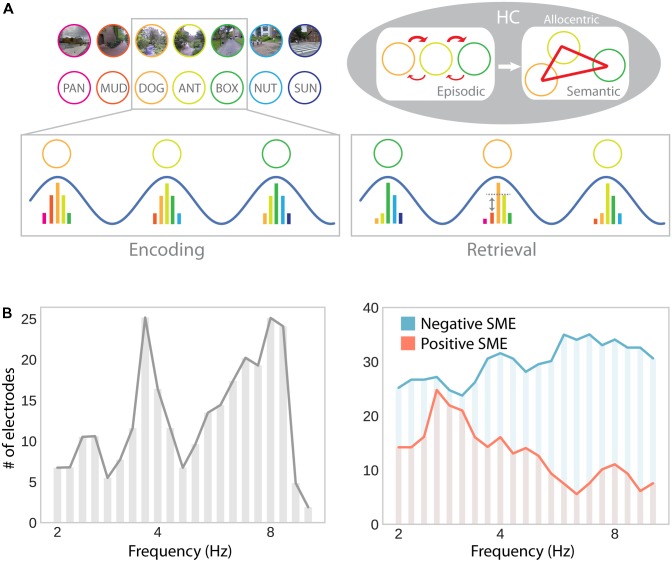
While extensive research on the neurophysiology of spatial memory has been carried out in rodents, memory research in humans had traditionally focused on more abstract, language-based tasks. Recent studies have begun to address this gap using virtual navigation tasks in combination with electrophysiological recordings in humans. These studies suggest that the human medial temporal lobe (MTL) is equipped with a population of place and grid cells similar to that previously observed in the rodent brain. Furthermore, theta oscillations have been linked to spatial navigation and, more specifically, to the encoding and retrieval of spatial information. While some studies suggest a single navigational theta rhythm which is of lower frequency in humans than rodents, other studies advocate for the existence of two functionally distinct delta-theta frequency bands involved in both spatial and episodic memory. Despite the general consensus between rodent and human electrophysiology, behavioral work in humans does not unequivocally support the use of a metric Euclidean map for navigation. Formal models of navigational behavior, which specifically consider the spatial scale of the environment and complementary learning mechanisms, may help to better understand different navigational strategies and their neurophysiological mechanisms. Finally, the functional overlap of spatial and declarative memory in the MTL calls for a unified theory of MTL function. Such a theory will critically rely upon linking task-related phenomena at multiple temporal and spatial scales. Understanding how single cell responses relate to ongoing theta oscillations during both the encoding and retrieval of spatial and non-spatial associations appears to be key toward developing a more mechanistic understanding of memory processes in the MTL.
Keywords: MTL; cognitive map; episodic memory; grid cells; navigation; place cells; spatial memory; theta.
Figures
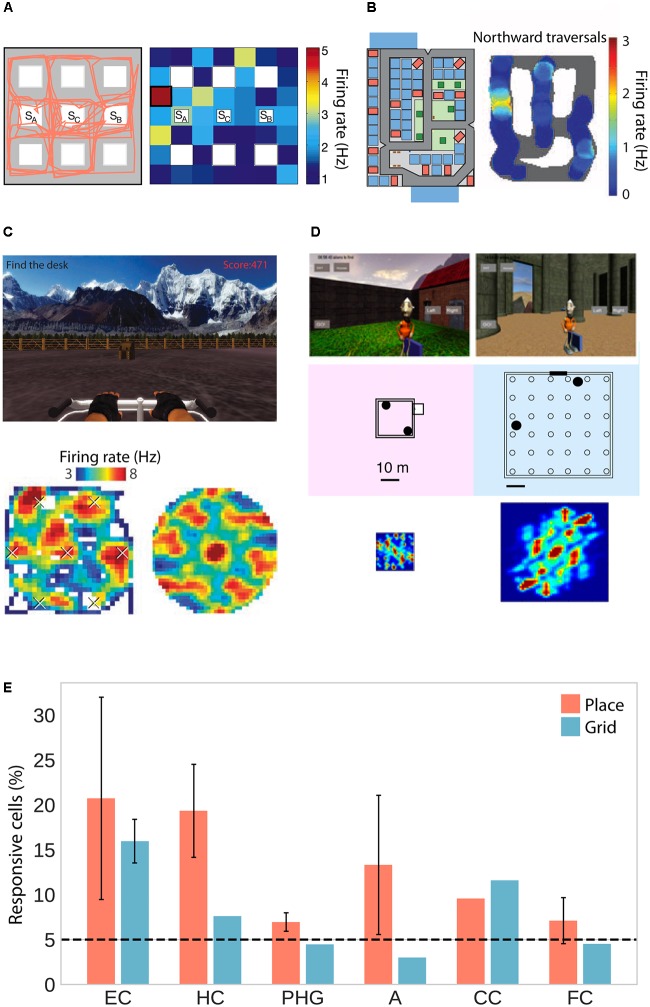
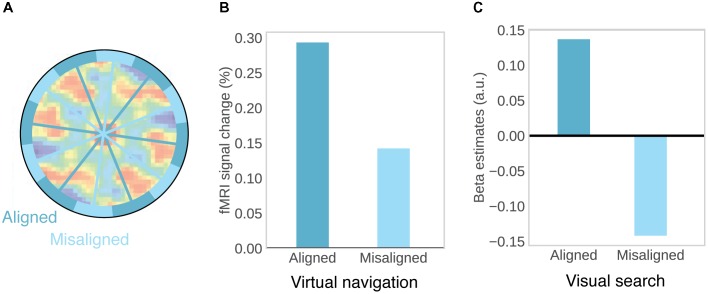

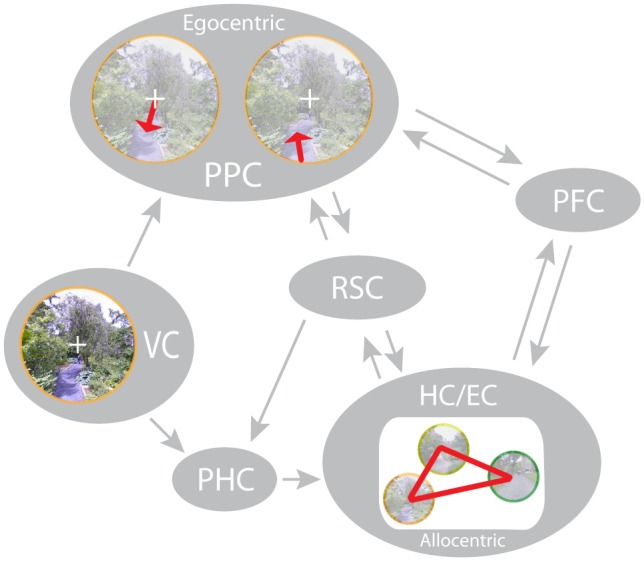
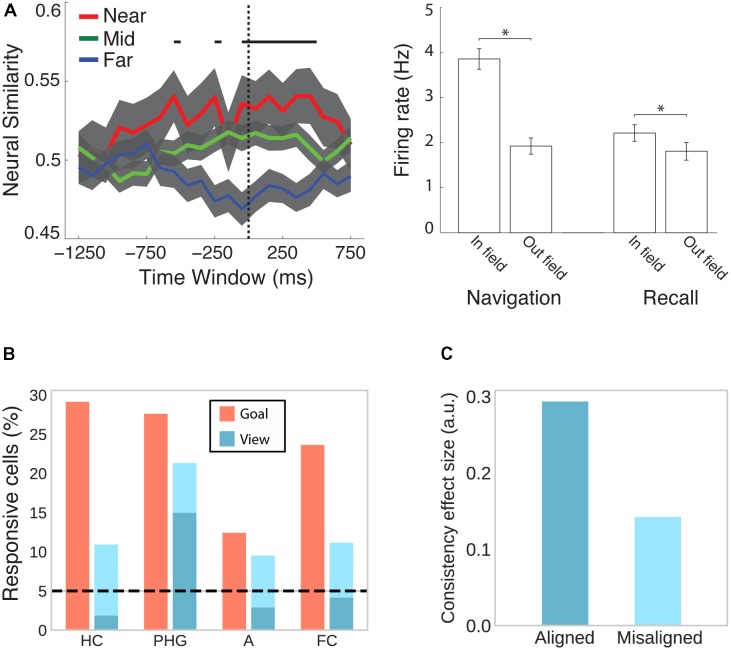
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous