

Prefrontal cortex modulates firing pattern in the nucleus reuniens of the midline thalamus via distinct corticothalamic pathways
- PMID: 30107061
- PMCID: PMC6237082
- DOI: 10.1111/ejn.14111
Prefrontal cortex modulates firing pattern in the nucleus reuniens of the midline thalamus via distinct corticothalamic pathways
Abstract
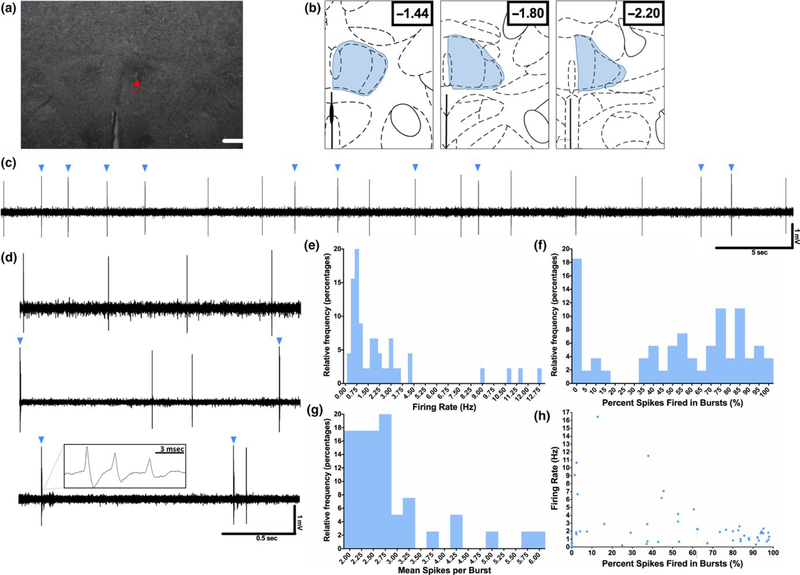
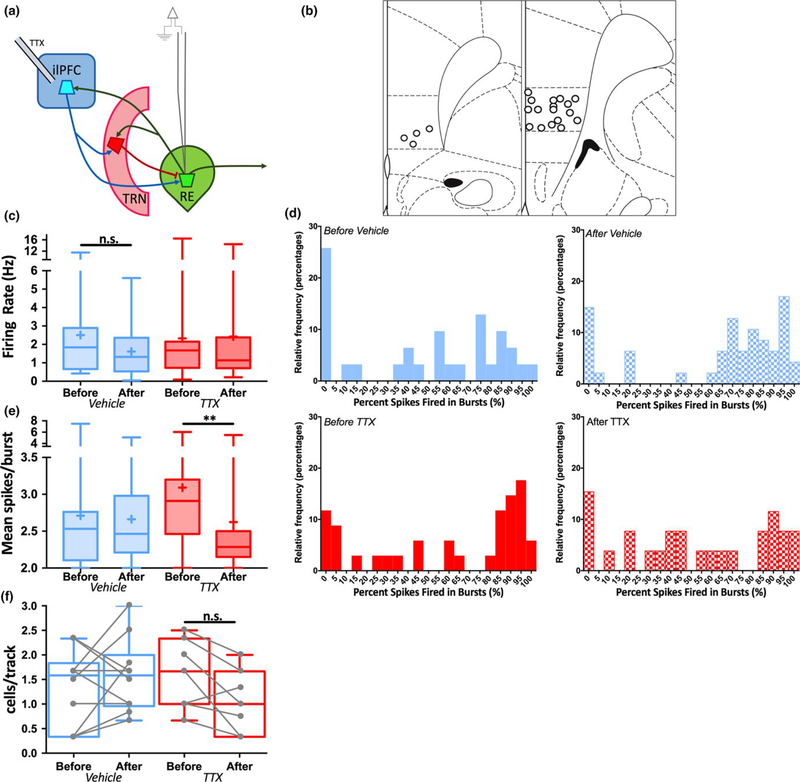
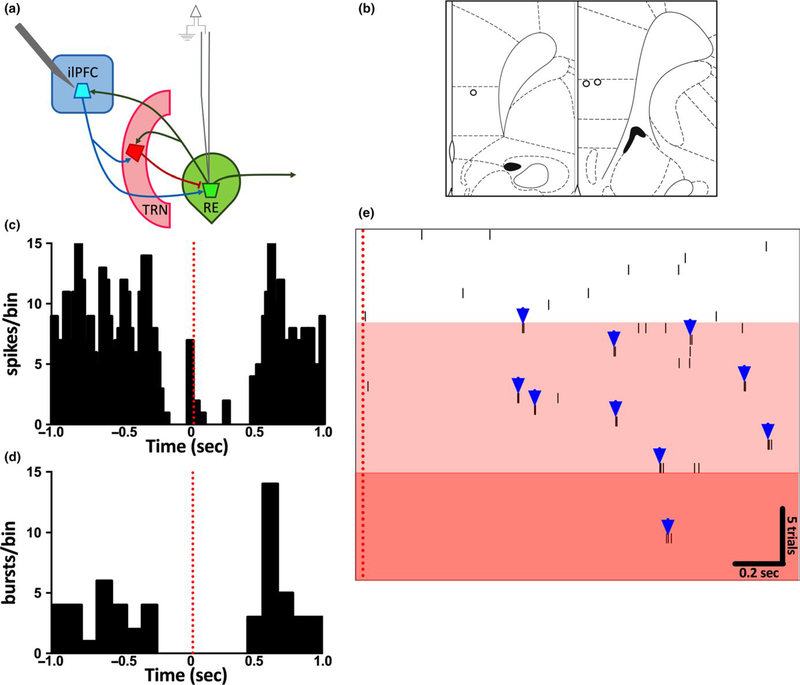
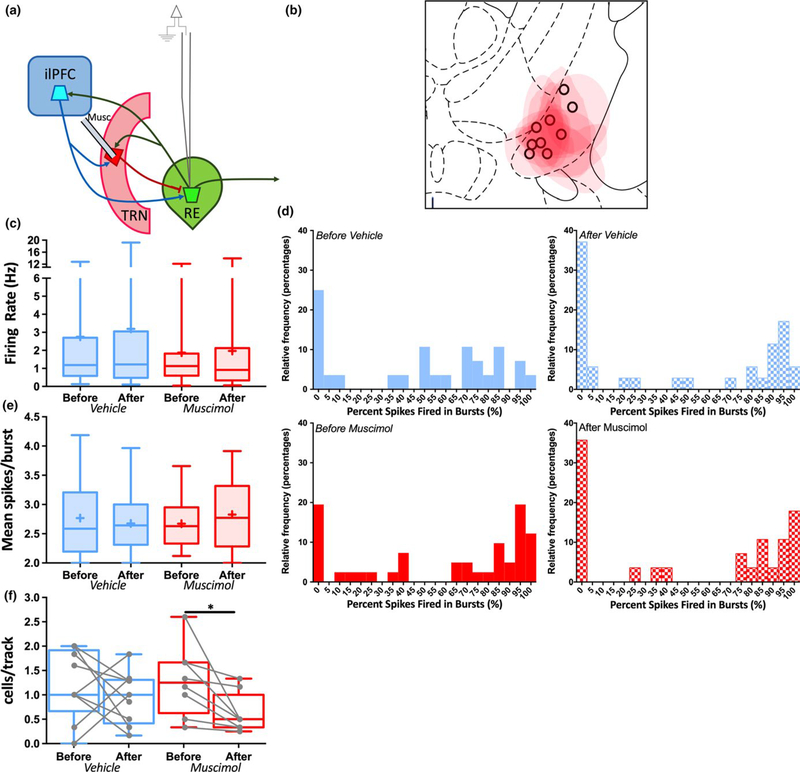
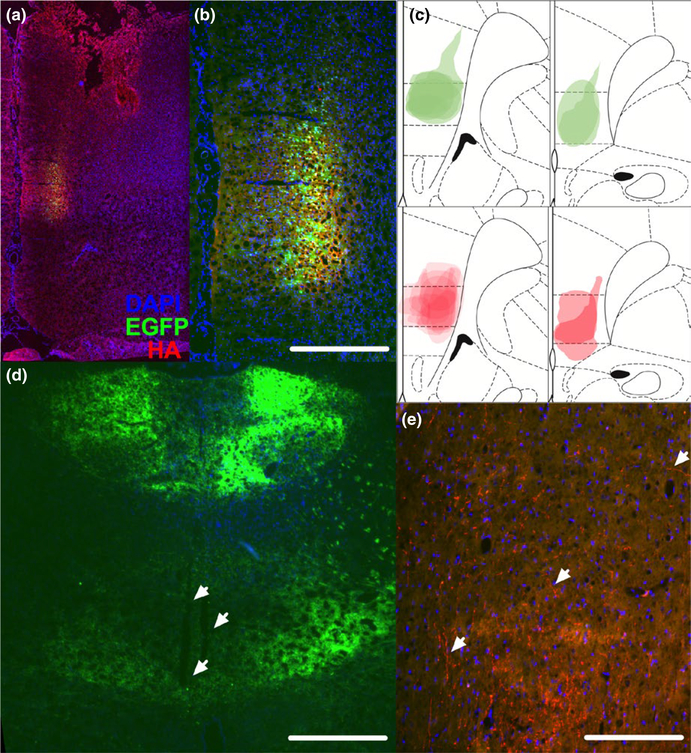
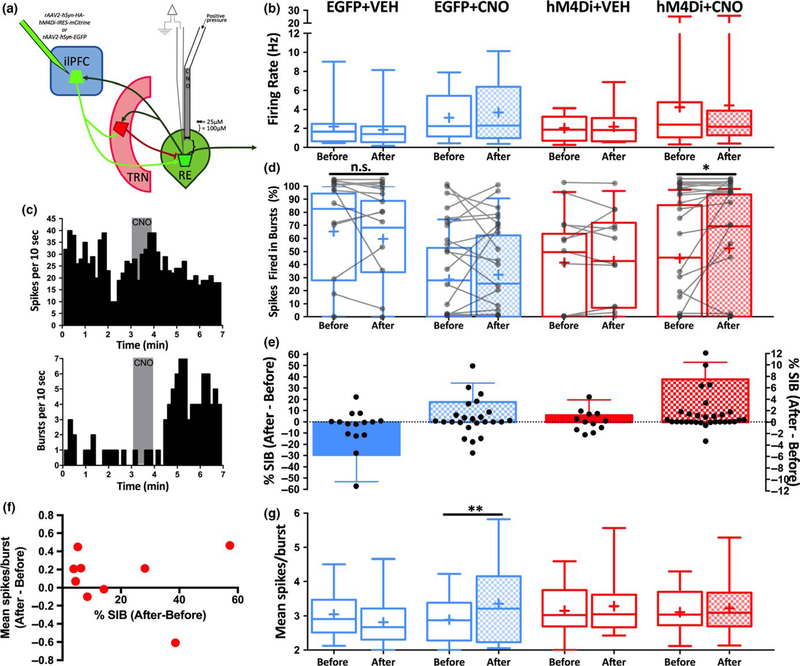
The thalamus has long been recognized for its role in relaying sensory information from the periphery, a function accomplished by its "first-order" nuclei. However, a second category of thalamic nuclei, termed "higher-order" nuclei, have been shown instead to mediate communication between cortical areas. The nucleus reuniens of the midline thalamus (RE) is a higher-order nucleus known to act as a conduit of reciprocal communication between the medial prefrontal cortex (mPFC) and hippocampus. While anatomical and behavioural studies of RE are numerous, circuit-based electrophysiological studies, particularly those examining the impact of cortical input and the thalamic reticular nucleus (TRN) on RE neuron firing, are sparse. To characterize RE neuron firing properties and dissect the circuit dynamics of the infralimbic subdivision of the mPFC (ilPFC), the TRN and RE, we used in vivo, extracellular, single-unit recordings in male Sprague Dawley rats and manipulated neural activity using targeted pharmacological manipulations, electrical stimulation and a projection-specific implementation of designer receptors exclusively activated by designer drugs (DREADDs). We show that ilPFC inhibition reduces multiple burst firing parameters in RE, whereas ilPFC stimulation drives burst firing and dampens tonic firing. In addition, TRN inhibition reduces the number of spontaneously active neurons in RE. Finally, inhibition of ilPFC terminals in RE selectively enhances a subset of burst firing parameters. These findings demonstrate that ilPFC input, both via direct projections and via the TRN, can modulate RE neuron firing pattern in nuanced and complex ways. They also highlight the ilPFC-TRN-RE circuit as a likely critical component of prefrontal-hippocampal interactions.
Keywords: corticothalamic circuits; limbic thalamus; thalamic bursting; thalamic reticular nucleus.
© 2018 Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Conflict of interest statement
CONFLICT OF INTEREST
ECZ declares no competing financial interests. AAG reports the following: Johnson & Johnson, Lundbeck, Pfizer, GSK, Merck, Takeda, Dainippon Sumitomo, Otsuka, Lilly, Roche, Asubio, Abbott, Autofony, Janssen, Alkermes, Newron.
Figures
Similar articles
-
The Nucleus Reuniens of the Midline Thalamus Gates Prefrontal-Hippocampal Modulation of Ventral Tegmental Area Dopamine Neuron Activity.J Neurosci. 2016 Aug 24;36(34):8977-84. doi: 10.1523/JNEUROSCI.1402-16.2016. J Neurosci. 2016. PMID: 27559178 Free PMC article.
-
Analysis of projections from the medial prefrontal cortex to the thalamus in the rat, with emphasis on nucleus reuniens.J Comp Neurol. 2002 Jan 7;442(2):163-87. doi: 10.1002/cne.10083. J Comp Neurol. 2002. PMID: 11754169
-
Excitatory actions of the ventral midline thalamus (rhomboid/reuniens) on the medial prefrontal cortex in the rat.Synapse. 2006 Jul;60(1):45-55. doi: 10.1002/syn.20271. Synapse. 2006. PMID: 16596625
-
The reuniens and rhomboid nuclei: neuroanatomy, electrophysiological characteristics and behavioral implications.Prog Neurobiol. 2013 Dec;111:34-52. doi: 10.1016/j.pneurobio.2013.08.006. Epub 2013 Sep 8. Prog Neurobiol. 2013. PMID: 24025745 Free PMC article. Review.
-
Limbic circuitry of the midline thalamus.Neurosci Biobehav Rev. 2015 Jul;54:89-107. doi: 10.1016/j.neubiorev.2015.01.014. Epub 2015 Jan 20. Neurosci Biobehav Rev. 2015. PMID: 25616182 Free PMC article. Review.
Cited by
-
Prefrontal cortex neurons encode ambient light intensity differentially across regions and layers.Nat Commun. 2024 Jun 29;15(1):5501. doi: 10.1038/s41467-024-49794-w. Nat Commun. 2024. PMID: 38951486 Free PMC article.
-
Isolated theta waves originating from the midline thalamus trigger memory reactivation during NREM sleep in mice.Nat Commun. 2024 Oct 25;15(1):9231. doi: 10.1038/s41467-024-53522-9. Nat Commun. 2024. PMID: 39455583 Free PMC article.
-
Dysregulation of Midbrain Dopamine System and the Pathophysiology of Schizophrenia.Front Psychiatry. 2020 Jun 30;11:613. doi: 10.3389/fpsyt.2020.00613. eCollection 2020. Front Psychiatry. 2020. PMID: 32719622 Free PMC article. Review.
-
Secondary thalamic neuroinflammation after focal cortical stroke and traumatic injury mirrors corticothalamic functional connectivity.J Comp Neurol. 2022 May;530(7):998-1019. doi: 10.1002/cne.25259. Epub 2021 Nov 1. J Comp Neurol. 2022. PMID: 34633669 Free PMC article.
-
The medial septum enhances reversal learning via opposing actions on ventral tegmental area and substantia nigra dopamine neurons.Neuropsychopharmacology. 2019 Dec;44(13):2186-2194. doi: 10.1038/s41386-019-0453-1. Epub 2019 Jul 1. Neuropsychopharmacology. 2019. PMID: 31261368 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
