

Interneuron Simplification and Loss of Structural Plasticity As Markers of Aging-Related Functional Decline
- PMID: 30108129
- PMCID: PMC6158697
- DOI: 10.1523/JNEUROSCI.0808-18.2018
Interneuron Simplification and Loss of Structural Plasticity As Markers of Aging-Related Functional Decline
Abstract
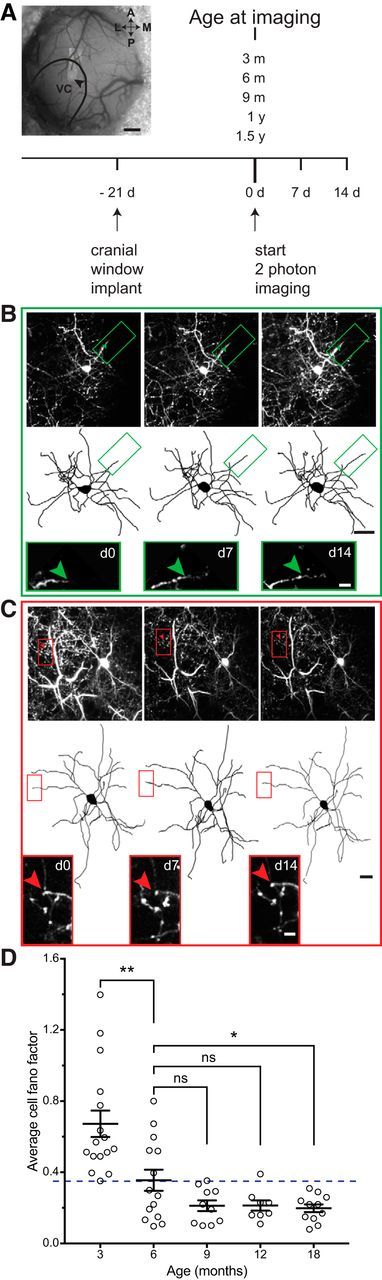
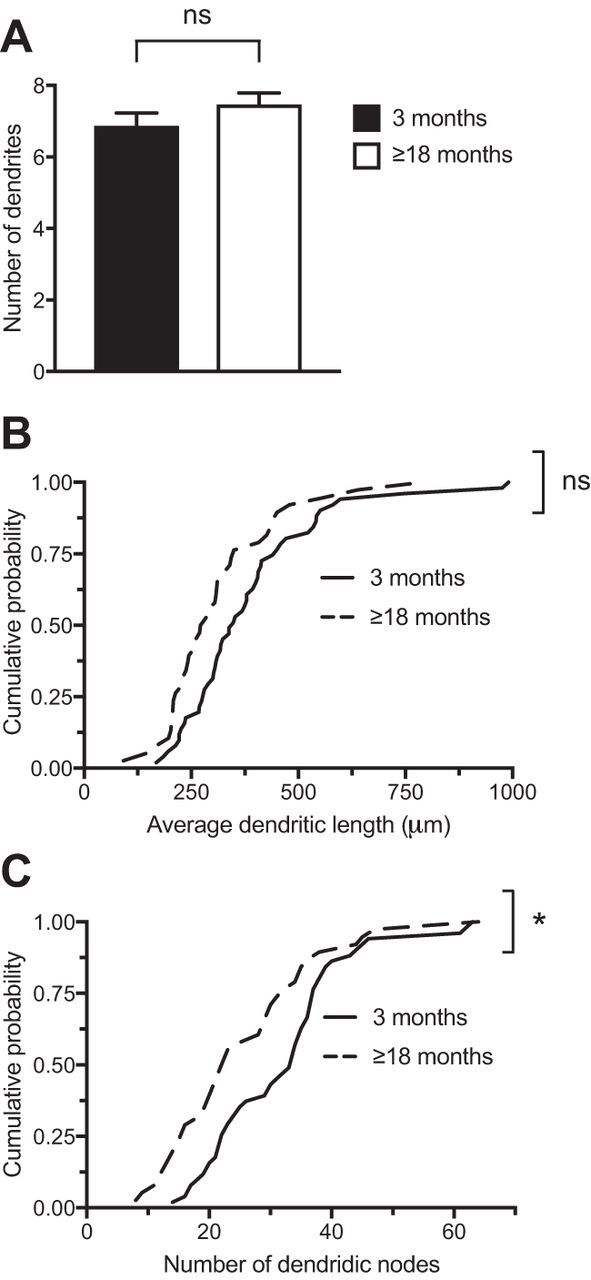
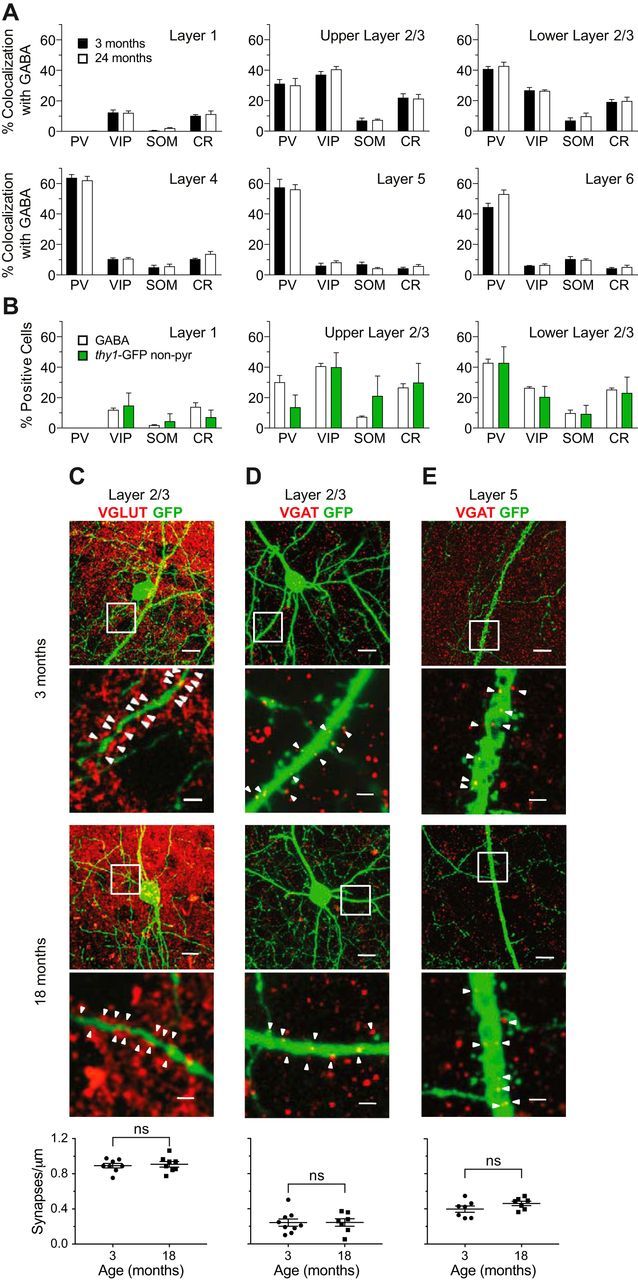
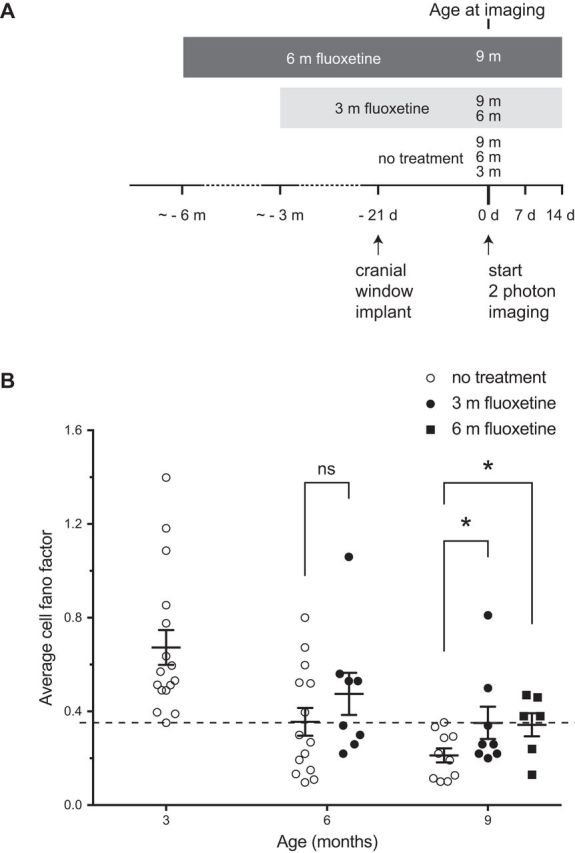
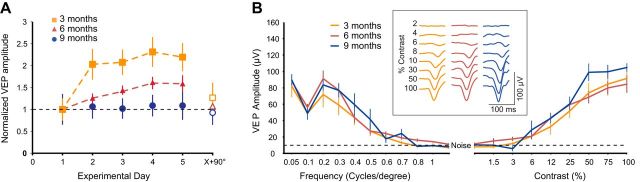
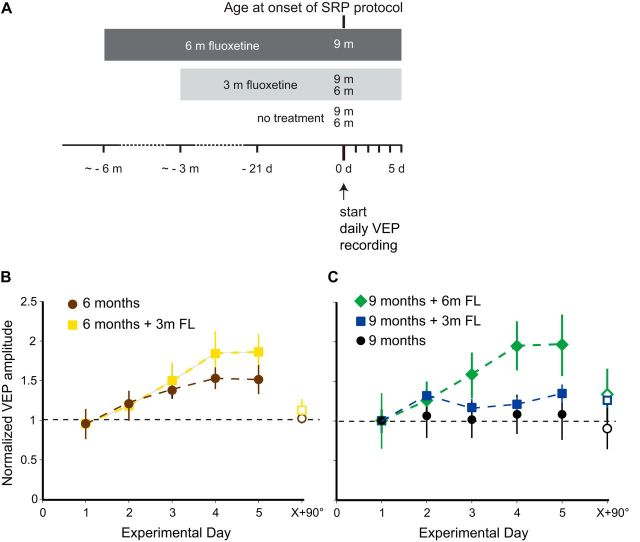
Changes in excitatory neuron and synapse structure have been recognized as a potential physical source of age-related cognitive decline. Despite the importance of inhibition to brain plasticity, little is known regarding aging-associated changes to inhibitory neurons. Here we test for age-related cellular and circuit changes to inhibitory neurons of mouse visual cortex. We find no substantial difference in inhibitory neuron number, inhibitory neuronal subtypes, or synapse numbers within the cerebral cortex of aged mice compared with younger adults. However, when comparing cortical interneuron morphological parameters, we find differences in complexity, suggesting that arbors are simplified in aged mice. In vivo two-photon microscopy has previously shown that in contrast to pyramidal neurons, inhibitory interneurons retain a capacity for dendritic remodeling in the adult. We find that this capacity diminishes with age and is accompanied by a shift in dynamics from balanced branch additions and retractions to progressive prevalence of retractions, culminating in a dendritic arbor that is both simpler and more stable. Recording of visually evoked potentials shows that aging-related interneuron dendritic arbor simplification and reduced dynamics go hand in hand with loss of induced stimulus-selective response potentiation (SRP), a paradigm for adult visual cortical plasticity. Chronic treatment with the antidepressant fluoxetine reversed deficits in interneuron structural dynamics and restored SRP in aged animals. Our results support a structural basis for age-related impairments in sensory perception, and suggest that declines in inhibitory neuron structural plasticity during aging contribute to reduced functional plasticity.SIGNIFICANCE STATEMENT Structural alterations in neuronal morphology and synaptic connections have been proposed as a potential physical basis for age-related decline in cognitive function. Little is known regarding aging-associated changes to inhibitory neurons, despite the importance of inhibitory circuitry to adult cortical plasticity and the reorganization of cortical maps. Here we show that brain aging goes hand in hand with progressive structural simplification and reduced plasticity of inhibitory neurons, and a parallel decline in sensory map plasticity. Fluoxetine treatment can attenuate the concurrent age-related declines in interneuron structural and functional plasticity, suggesting it could provide an important therapeutic approach for mitigating sensory and cognitive deficits associated with aging.
Keywords: aging; fluoxetine; inhibitory neurons; mice; two-photon microscopy; visually evoked potential.
Copyright © 2018 the authors 0270-6474/18/388421-12$15.00/0.
Figures
References
-
- Chattopadhyaya B, Di Cristo G, Higashiyama H, Knott GW, Kuhlman SJ, Welker E, Huang ZJ (2004) Experience and activity-dependent maturation of perisomatic GABAergic innervation in primary visual cortex during a postnatal critical period. J Neurosci 24:9598–9611. 10.1523/JNEUROSCI.1851-04.2004 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials