

Review
doi: 10.1152/physiol.00025.2018.
Defining the Rhythmogenic Elements of Mammalian Breathing
Affiliations
- PMID: 30109823
- PMCID: PMC6230551
- DOI: 10.1152/physiol.00025.2018
Item in Clipboard
Review
Defining the Rhythmogenic Elements of Mammalian Breathing
Physiology (Bethesda).
.
Abstract
Breathing's remarkable ability to adapt to changes in metabolic, environmental, and behavioral demands stems from a complex integration of its rhythm-generating network within the wider nervous system. Yet, this integration complicates identification of its specific rhythmogenic elements. Based on principles learned from smaller rhythmic networks of invertebrates, we define criteria that identify rhythmogenic elements of the mammalian breathing network and discuss how they interact to produce robust, dynamic breathing.
Figures
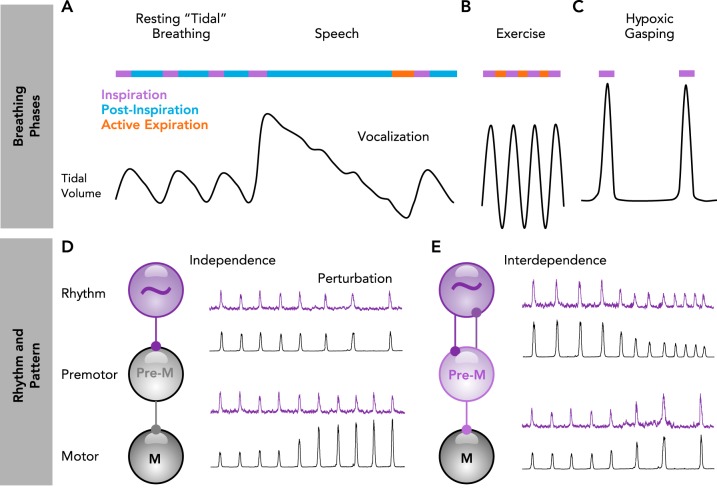
Breathing phases arise from a combination of rhythm- and pattern-generating mechanisms A–C: schematic of breathing phase coordination and reconfiguration during rest, speech, exercise, and severe hypoxia. A: during rest, breathing alternates between inspiration (purple) and postinspiration (blue). Prior to speaking, the depth of inspiration (tidal volume) is adjusted to the anticipated length of the vocalization. Vocalization occurs during an extended postinspiratory phase. B: during heavy exercise, breathing generally alternates rapidly between inspiration and active expiration (red) to match increased metabolic demands. C: during severe hypoxia, breathing reconfigures to produce a slow one-phase inspiratory rhythm. D–E: processes of rhythm and pattern generation can be independent or interdependent. D: when independent, perturbations of rhythm (purple) do not alter pattern (black), and perturbations of pattern do not alter rhythm. E: when interdependent, perturbations of rhythm also alter pattern, and perturbations of pattern also alter rhythm.
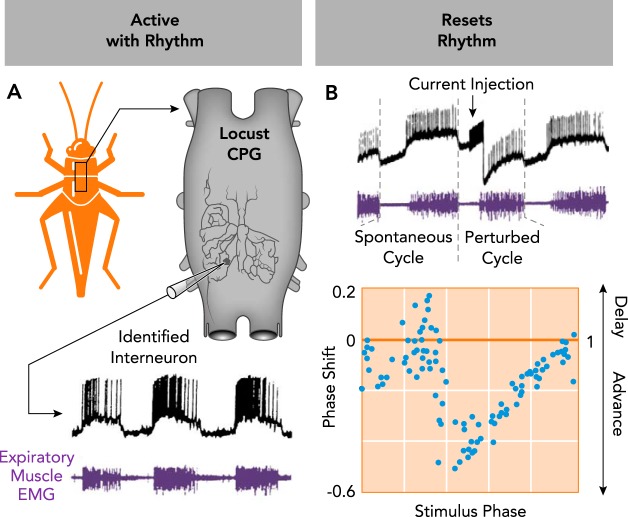
Characteristics of an identified rhythmogenic neuron in the respiratory central pattern generator of the locust A: the neuron (subesophogeal ganglion interneuron 378) is active in phase with the rhythm (criterion 1), as indicated by intracellular recording of spiking activity (black) concurrent with expiratory muscle EMG activity (purple). B: stimulation of the neuron resets the respiratory rhythm (criterion 2). Direct current stimulation of subesophogeal ganglion interneuron 378 shortens the perturbed respiratory cycle relative to a spontaneous cycle. Resetting characteristics for the neuron are visualized in a phase-shift plot, demonstrating stimulus phase-dependent resetting of the rhythm. Data pooled from experiments in three different animals. A and B are adapted from Ref. with permission from the Journal of Neurophysiology.
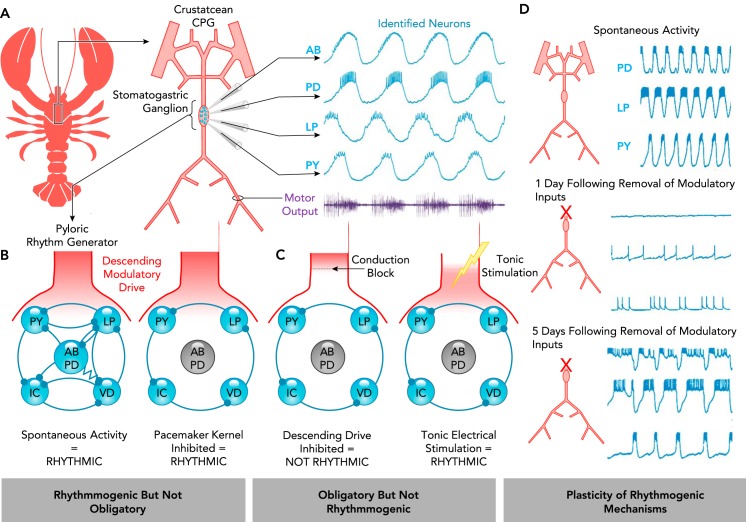
The rhythmogenic role of an element may not be determined solely based on whether it is obligatory A: schematic of the crustacean stomatogastric nervous system and intracellular recordings of identified stomatogastric ganglion neurons of the pyloric rhythm generator (AB, anterior burster; PD, pyloric dilator; LP, lateral pyloric; PY, pyloric). The electrically coupled AB and PD neurons have autonomous bursting properties and comprise the “pacemaker kernel.” These neurons are active in phase with rhythmic motor output (purple) and can also reset the rhythm. Adapted from Ref. , with permission from the Annual Review of Physiology. B–C: schematic of the pyloric rhythm-generating circuit. All synapses are inhibitory; resistor indicates electrical connections. B: in the presence of descending neuromodulatory drive, silencing the AB/PD pacemaker kernel does not abolish rhythmogenesis. C: subsequent removal of neuromodulatory drive eliminates the rhythm; however, it can be restored with artificial tonic stimulation (129). D: the extent to which a given mechanism is obligatory for rhythmogenesis may be altered by plasticity. Spontaneous rhythmicity of the pyloric rhythm generator is initially lost following removal of neuromodulatory inputs. However, rhythmicity returns as the rhythm-generating properties of the network adapt. Adapted from Ref. , with permission from the Journal of Neurophysiology.
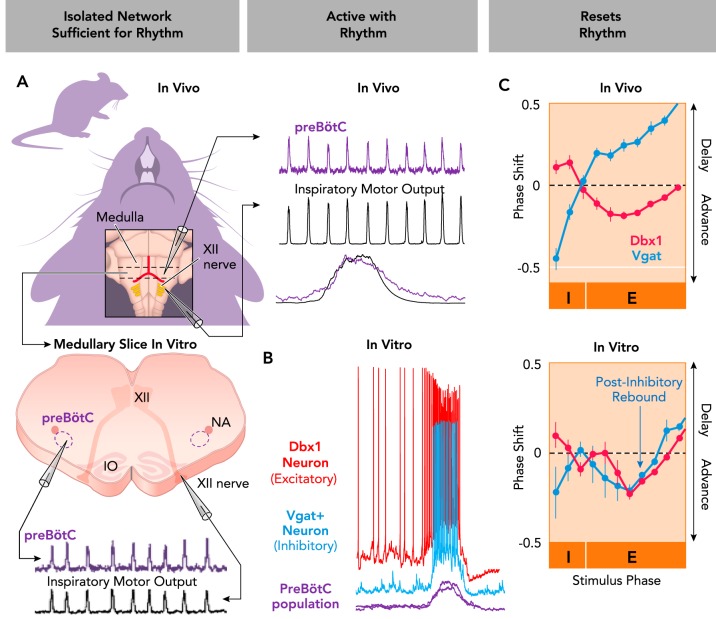
Rhythmogenic roles of excitatory and inhibitory neurons in the mammalian breathing CPG A: the pre-Bötzinger Complex (preBötC) continues to produce a rhythm (purple) that is in phase with inspiratory hypoglossal (XII) motor output (black) when isolated in medullary slices from neonatal mice. The preBötC is also active in phase with inspiratory motor activity in intact adult mice. B: excitatory (red) and inhibitory (blue) preBötC neurons are concurrently active in phase with rhythmic inspiratory activity (purple). Intracellular current-clamp recordings of genetically identified Dbx1+ (excitatory) and Vgat+ (inhibitory) neurons active with integrated extracellular preBötC population bursts generated in vitro. C: optogenetic stimulation of excitatory (Dbx1+) or inhibitory (Vgat+) preBötC neurons resets the breathing rhythm in vivo and in vitro. Stimulations can advance or delay the next inspiratory cycle depending on stimulus phase (I, inspiratory phase; E, expiratory phase). Note that postinhibitory rebound advances the next cycle during relatively slow rhythms in vitro but not in vivo. Figure adapted from Ref. , with permission from Nature Communications.
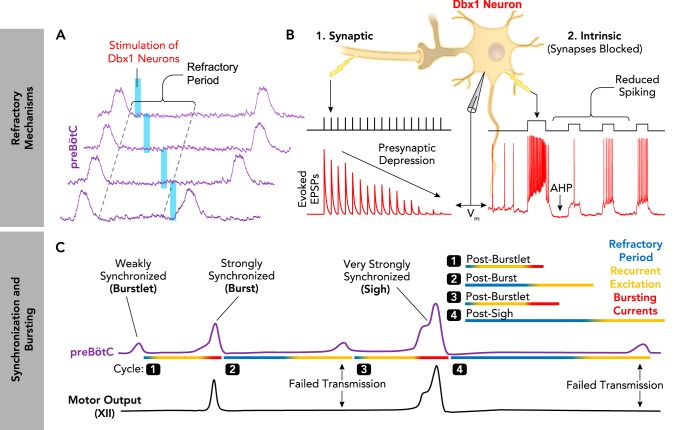
The relationship between synchronization, bursting, and refractory mechanisms in the preBötC A: the preBötC has an inherent refractoriness. The probability of evoking preBötC population bursts by optogenetic stimulation of Dbx1 neurons is transiently reduced following spontaneous preBötC bursts, i.e., during the refractory period (9) B: synaptic and intrinsic properties of excitatory Dbx1+ neurons contribute to the refractory period (1). Schematic demonstrating presynaptic depression of excitatory synaptic transmission between preBötC neurons. The amplitude of evoked excitatory postsynaptic potentials (EPSPs; red) recorded in Dbx1 neurons is progressively reduced during repeated electrical stimulations (black) of presynaptic inputs (62). 2: intrinsic properties transiently reduce the excitability of Dbx1 neurons following preBötC bursts. Intracellular current-clamp recording of a synaptically isolated Dbx1 neuron demonstrating membrane afterhyperpolarization (AHP) and transiently reduced spiking following an initial, larger depolarizing burst (9). C: schematic representing the different amounts of synchronization during distinct patterns of preBötC activity (purple) and associated hypoglossal (XII) motor output (black). During burstlets, synchronization is weak, bursting currents are only activated in some neurons, and the refractory period is minimal. During normal “eupneic” bursts, synchronization is strong and bursting currents are activated in many neurons, resulting in a period of relative refractoriness. During periodic sigh bursts, synchronization is very strong and bursting currents are strongly activated, leading to large depolarizing drive potentials in preBötC neurons and an exaggerated refractory period. Note that strong synchronization facilitated by activation of bursting currents promotes successful transmission of preBötC activity to XII motor output.
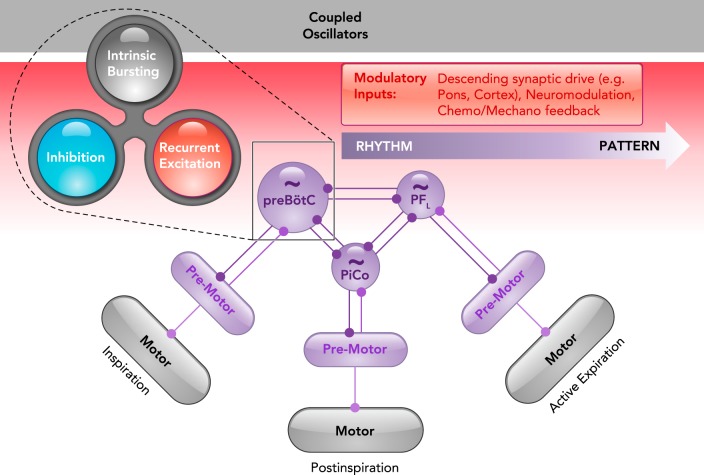
A contemporary view on the origins of mammalian breathing The triple oscillator hypothesis (6) proposes that each breathing phase is generated by a distinct microcircuit in the medulla: the preBötzinger complex (preBötC) generates inspiration, the postinspiratory complex (PiCo) generates postinspiration, and the lateral parafacial region (pFL) generates active expiration. Each oscillator is coupled by excitatory and inhibitory connections, although inhibition typically dominates to coordinate the timing of each breathing phase. As shown in the preBötC, rhythmicity within each microcircuit is controlled by a balance between recurrent synaptic excitation, inhibition, and intrinsic bursting properties, i.e., the rhythmogenic triangle (103). A gradient of rhythm (purple) and pattern (gray) generating properties can be independent or interdependent depending on the connectivity between specific rhythm, premotor, and motor elements. The activity of rhythm- and pattern-generating elements can be differentially tuned by various modulatory inputs (red) to endow breathing with exquisite metabolic-, state-, and behavior-dependent control.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources