

A unified computational framework for modeling genome-wide nucleosome landscape
- PMID: 30113318
- PMCID: PMC6170202
- DOI: 10.1088/1478-3975/aadad2
A unified computational framework for modeling genome-wide nucleosome landscape
Abstract
Nucleosomes form the fundamental building blocks of eukaryotic chromatin, and previous attempts to understand the principles governing their genome-wide distribution have spurred much interest and debate in biology. In particular, the precise role of DNA sequence in shaping local chromatin structure has been controversial. This paper rigorously quantifies the contribution of hitherto-debated sequence features-including G+C content, 10.5 bp periodicity, and poly(dA:dT) tracts-to three distinct aspects of genome-wide nucleosome landscape: occupancy, translational positioning and rotational positioning. Our computational framework simultaneously learns nucleosome number and nucleosome-positioning energy from genome-wide nucleosome maps. In contrast to other previous studies, our model can predict both in vitro and in vivo nucleosome maps in Saccharomyces cerevisiae. We find that although G+C content is the primary determinant of MNase-derived nucleosome occupancy, MNase digestion biases may substantially influence this GC dependence. By contrast, poly(dA:dT) tracts are seen to deter nucleosome formation, regardless of the experimental method used. We further show that the 10.5 bp nucleotide periodicity facilitates rotational but not translational positioning. Applying our method to in vivo nucleosome maps demonstrates that, for a subset of genes, the regularly-spaced nucleosome arrays observed around transcription start sites can be partially recapitulated by DNA sequence alone. Finally, in vivo nucleosome occupancy derived from MNase-seq experiments around transcription termination sites can be mostly explained by the genomic sequence. Implications of these results and potential extensions of the proposed computational framework are discussed.
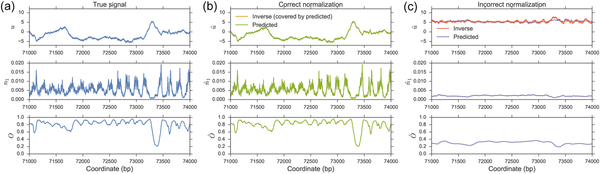
Figures






References
-
- Hughes AL and Rando OJ, “Mechanisms underlying nucleosome positioning in vivo,” Annual review of biophysics, vol. 43, pp. 41–63, 2014. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous