

Immobility in the sedentary plant-parasitic nematode H. glycines is associated with remodeling of neuromuscular tissue
- PMID: 30114260
- PMCID: PMC6095618
- DOI: 10.1371/journal.ppat.1007198
Immobility in the sedentary plant-parasitic nematode H. glycines is associated with remodeling of neuromuscular tissue
Abstract
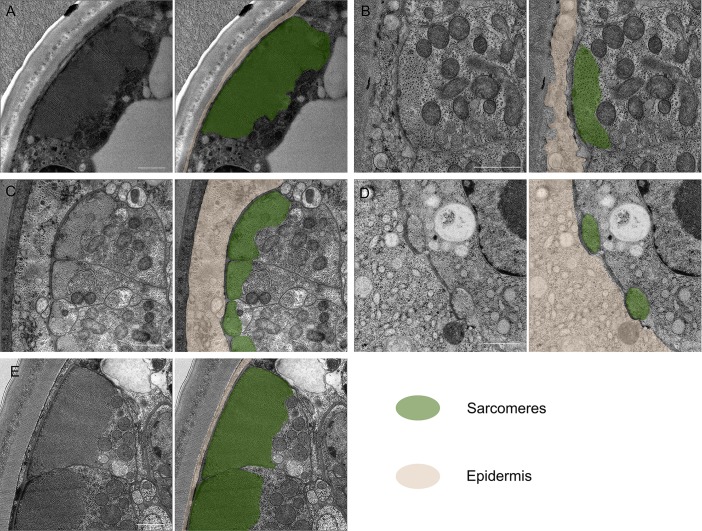
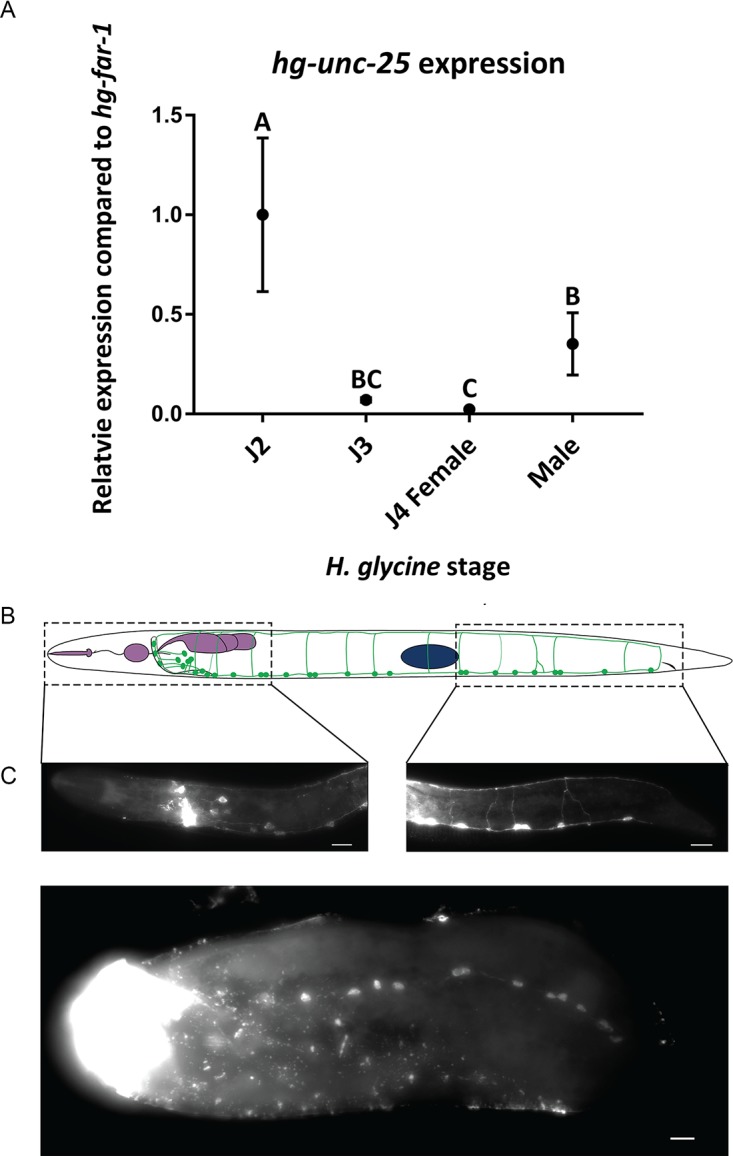
The sedentary plant-parasitic nematodes are considered among the most economically damaging pathogens of plants. Following infection and the establishment of a feeding site, sedentary nematodes become immobile. Loss of mobility is reversed in adult males while females never regain mobility. The structural basis for this change in mobility is unknown. We used a combination of light and transmission electron microscopy to demonstrate cell-specific muscle atrophy and sex-specific renewal of neuromuscular tissue in the sedentary nematode Heterodera glycines. We found that both females and males undergo body wall muscle atrophy and loss of attachment to the underlying cuticle during immobile developmental stages. Male H. glycines undergo somatic muscle renewal prior to molting into a mobile adult. In addition, we found developmental changes to the organization and number of motor neurons in the ventral nerve cord correlated with changes in mobility. To further examine neuronal changes associated with immobility, we used a combination of immunohistochemistry and molecular biology to characterize the GABAergic nervous system of H. glycines during mobile and immobile stages. We cloned and confirmed the function of the putative H. glycines GABA synthesis-encoding gene hg-unc-25 using heterologous rescue in C. elegans. We found a reduction in gene expression of hg-unc-25 as well as a reduction in the number of GABA-immunoreactive neurons during immobile developmental stages. Finally, we found evidence of similar muscle atrophy in the phylogenetically diverged plant-parasitic nematode Meloidogyne incognita. Together, our data demonstrate remodeling of neuromuscular structure and function during sedentary plant-parasitic nematode development.
Conflict of interest statement
The authors have declared no competing interests exist.
Figures






Similar articles
-
A decline in transcript abundance for Heterodera glycines homologs of Caenorhabditis elegans uncoordinated genes accompanies its sedentary parasitic phase.BMC Dev Biol. 2007 Apr 19;7:35. doi: 10.1186/1471-213X-7-35. BMC Dev Biol. 2007. PMID: 17445261 Free PMC article.
-
Embryogenesis in the parasitic nematode Heterodera glycines is independent of host-derived hatching stimulation.BMC Dev Biol. 2017 Jan 11;17(1):2. doi: 10.1186/s12861-016-0144-7. BMC Dev Biol. 2017. PMID: 28077087 Free PMC article.
-
The Heterodera glycines effector Hg16B09 is required for nematode parasitism and suppresses plant defense response.Plant Sci. 2019 Dec;289:110271. doi: 10.1016/j.plantsci.2019.110271. Epub 2019 Sep 14. Plant Sci. 2019. PMID: 31623793
-
Advances in Understanding the Molecular Mechanisms of Root Lesion Nematode Host Interactions.Annu Rev Phytopathol. 2016 Aug 4;54:253-78. doi: 10.1146/annurev-phyto-080615-100257. Epub 2016 Jan 1. Annu Rev Phytopathol. 2016. PMID: 27296144 Review.
-
Root-knot nematodes manipulate plant cell functions during a compatible interaction.J Plant Physiol. 2008 Jan;165(1):104-13. doi: 10.1016/j.jplph.2007.05.007. Epub 2007 Aug 6. J Plant Physiol. 2008. PMID: 17681399 Review.
Cited by
-
Whole mount multiplexed visualization of DNA, mRNA, and protein in plant-parasitic nematodes.Plant Methods. 2023 Dec 4;19(1):139. doi: 10.1186/s13007-023-01112-z. Plant Methods. 2023. PMID: 38049899 Free PMC article.
-
Convergent evolution of saccate body shapes in nematodes through distinct developmental mechanisms.Evodevo. 2019 Mar 14;10:5. doi: 10.1186/s13227-019-0118-5. eCollection 2019. Evodevo. 2019. PMID: 30911368 Free PMC article.
-
FMRFamide-Like Peptide 22 Influences the Head Movement, Host Finding, and Infection of Heterodera glycines.Front Plant Sci. 2021 Jun 22;12:673354. doi: 10.3389/fpls.2021.673354. eCollection 2021. Front Plant Sci. 2021. PMID: 34239524 Free PMC article.
-
Electron Microscopy Reveals Unexpected Cytoplasm and Envelope Changes during Thymineless Death in Escherichia coli.J Bacteriol. 2021 Aug 9;203(17):e0015021. doi: 10.1128/JB.00150-21. Epub 2021 Aug 9. J Bacteriol. 2021. PMID: 34152201 Free PMC article.
-
GABA Immunoreactivity and Pharmacological Effects vary Among Stylet-Bearing Nematodes.J Nematol. 2023 Nov 18;55(1):20230049. doi: 10.2478/jofnem-2023-0049. eCollection 2023 Feb. J Nematol. 2023. PMID: 38026555 Free PMC article.
References
-
- Koenning SR, Wrather JA. Suppression of soybean yield potential in the continental United States by plant diseases from 2006 to 2009. Plant Heal Prog. 2010; 10.1094/PHP-2010-1122-01-RS - DOI
-
- Wyss U, Zunke U. Observation of the behaviour of second stage juveniles of Heterodera schachtii inside host roots. Rev Nematol. 1986;9: 153–166.
-
- Elsea JR. The histological anatomy of the nematode Meloidogyne hapla (Heteroderidae). Proc Helm Soc Wash. 1951;18: 53–63.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
