

The Muscle Fiber Profiles, Mitochondrial Content, and Enzyme Activities of the Exceptionally Well-Trained Arm and Leg Muscles of Elite Cross-Country Skiers
- PMID: 30116201
- PMCID: PMC6084043
- DOI: 10.3389/fphys.2018.01031
The Muscle Fiber Profiles, Mitochondrial Content, and Enzyme Activities of the Exceptionally Well-Trained Arm and Leg Muscles of Elite Cross-Country Skiers
Abstract
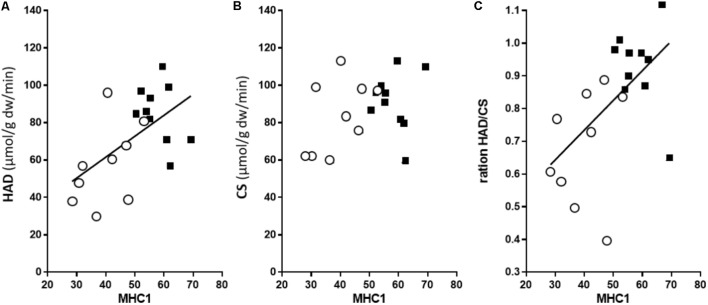
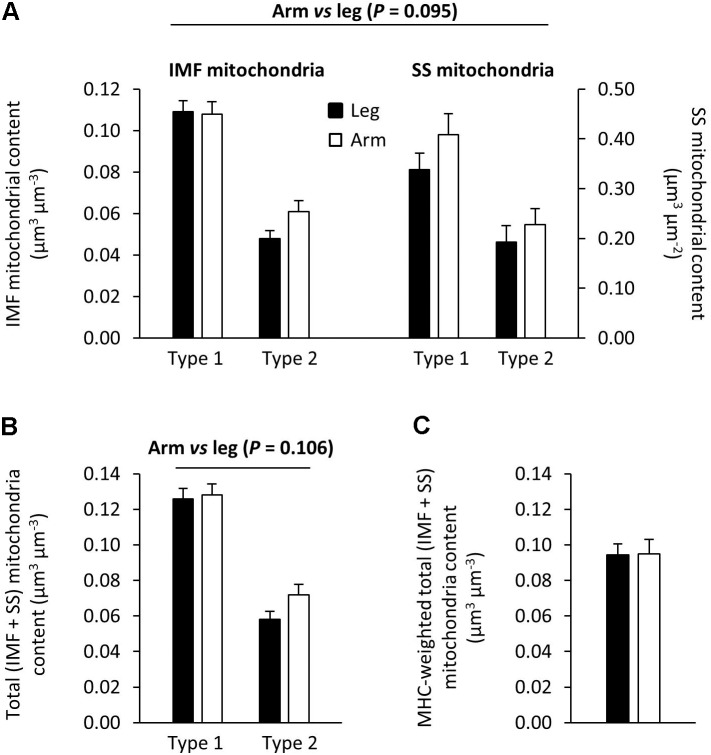
As one of the most physically demanding sports in the Olympic Games, cross-country skiing poses considerable challenges with respect to both force generation and endurance during the combined upper- and lower-body effort of varying intensity and duration. The isoforms of myosin in skeletal muscle have long been considered not only to define the contractile properties, but also to determine metabolic capacities. The current investigation was designed to explore the relationship between these isoforms and metabolic profiles in the arms (triceps brachii) and legs (vastus lateralis) as well as the range of training responses in the muscle fibers of elite cross-country skiers with equally and exceptionally well-trained upper and lower bodies. The proportion of myosin heavy chain (MHC)-1 was higher in the leg (58 ± 2% [34-69%]) than arm (40 ± 3% [24-57%]), although the mitochondrial volume percentages [8.6 ± 1.6 (leg) and 9.0 ± 2.0 (arm)], and average number of capillaries per fiber [5.8 ± 0.8 (leg) and 6.3 ± 0.3 (arm)] were the same. In these comparable highly trained leg and arm muscles, the maximal citrate synthase (CS) activity was the same. Still, 3-hydroxy-acyl-CoA-dehydrogenase (HAD) capacity was 52% higher (P < 0.05) in the leg compared to arm muscles, suggesting a relatively higher capacity for lipid oxidation in leg muscle, which cannot be explained by the different fiber type distributions. For both limbs combined, HAD activity was correlated with the content of MHC-1 (r2 = 0.32, P = 0.011), whereas CS activity was not. Thus, in these highly trained cross-country skiers capillarization of and mitochondrial volume in type 2 fiber can be at least as high as in type 1 fibers, indicating a divergence between fiber type pattern and aerobic metabolic capacity. The considerable variability in oxidative metabolism with similar MHC profiles provides a new perspective on exercise training. Furthermore, the clear differences between equally well-trained arm and leg muscles regarding HAD activity cannot be explained by training status or MHC distribution, thereby indicating an intrinsic metabolic difference between the upper and lower body. Moreover, trained type 1 and type 2A muscle fibers exhibited similar aerobic capacity regardless of whether they were located in an arm or leg muscle.
Keywords: IMCL; capillarization; cross-country skiing; fiber plasticity; limb muscles; mitochondria; training.
Figures

References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials