

Phylogeny and genetic diversity of the banana Fusarium wilt pathogen Fusarium oxysporum f. sp. cubense in the Indonesian centre of origin
- PMID: 30122796
- PMCID: PMC6086327
- DOI: 10.1016/j.simyco.2018.06.003
Phylogeny and genetic diversity of the banana Fusarium wilt pathogen Fusarium oxysporum f. sp. cubense in the Indonesian centre of origin
Abstract
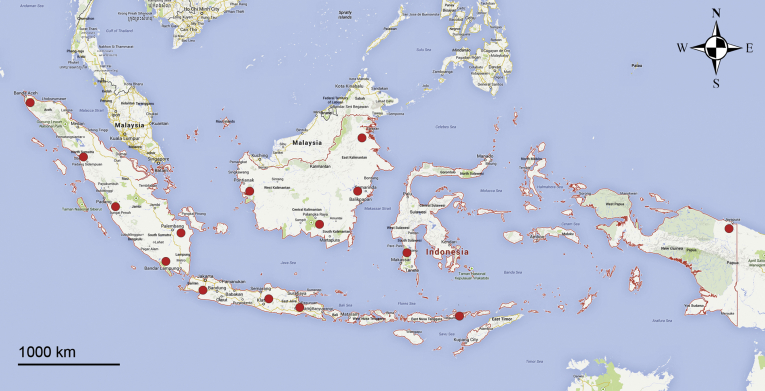
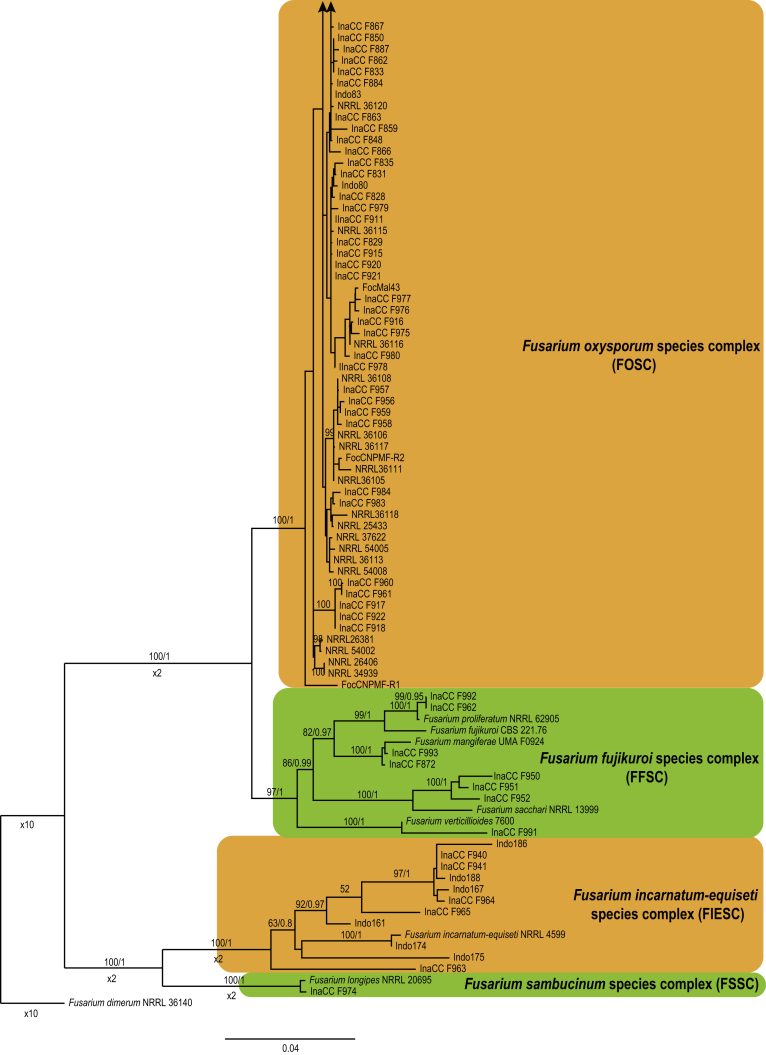
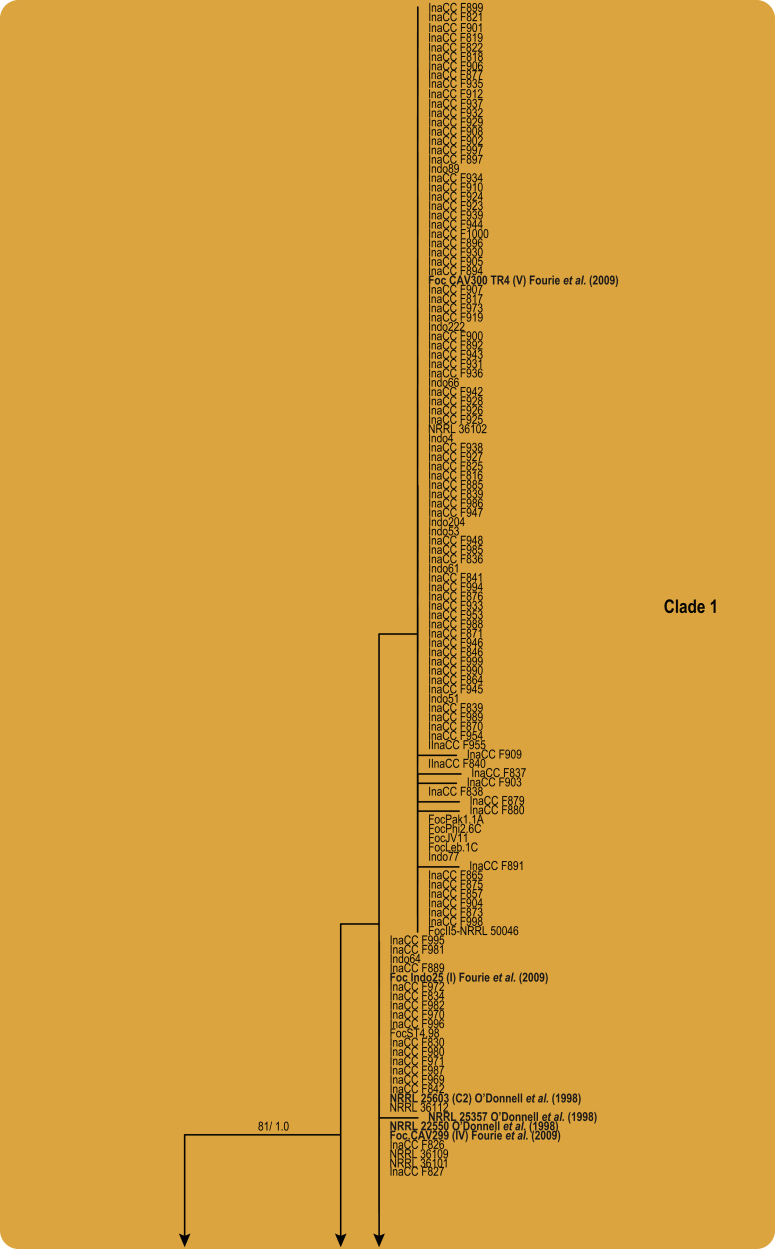
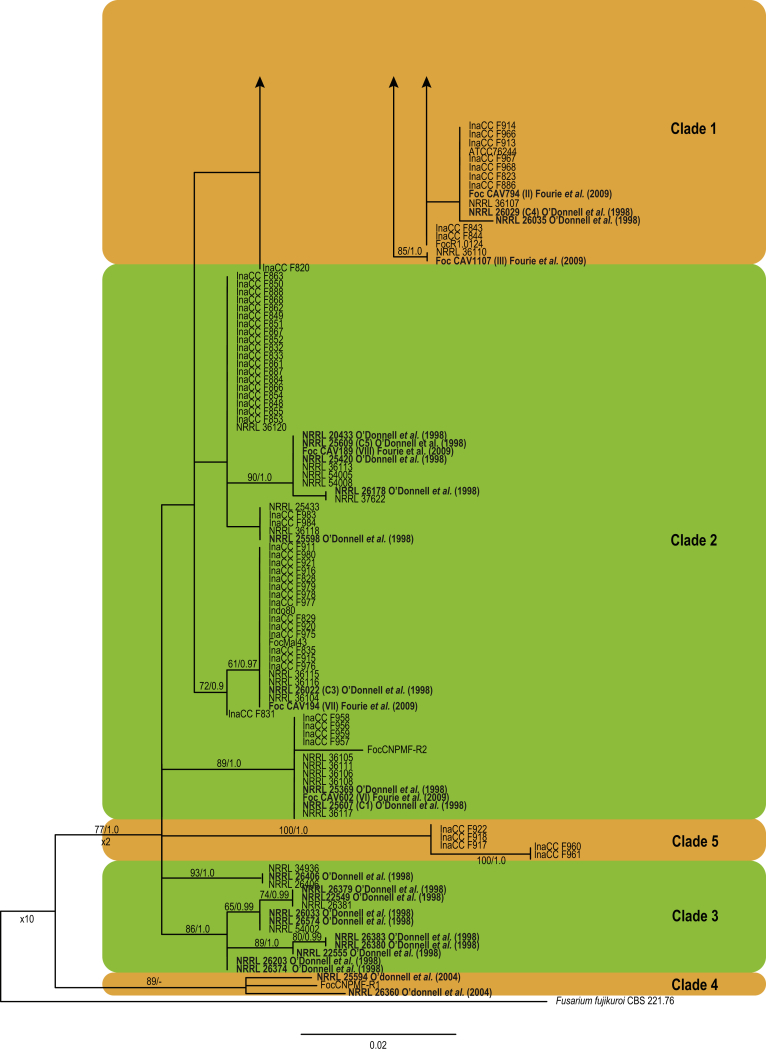
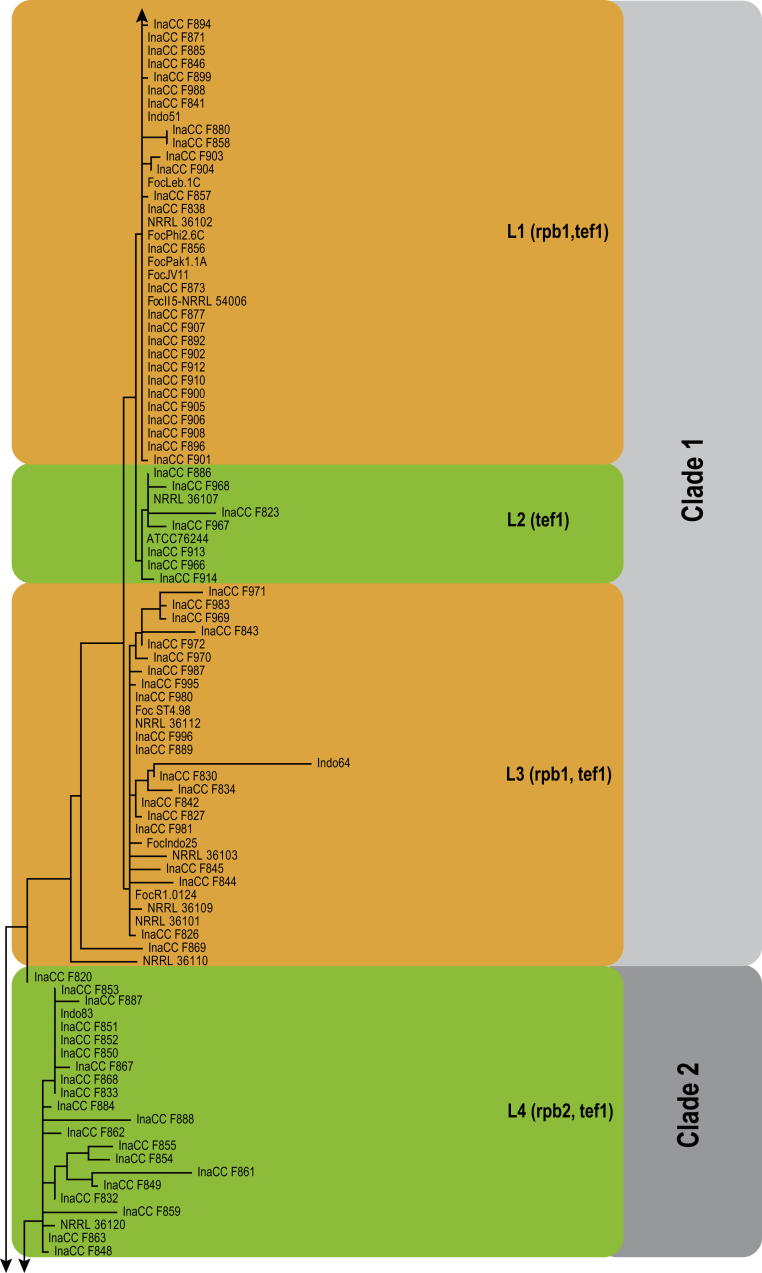
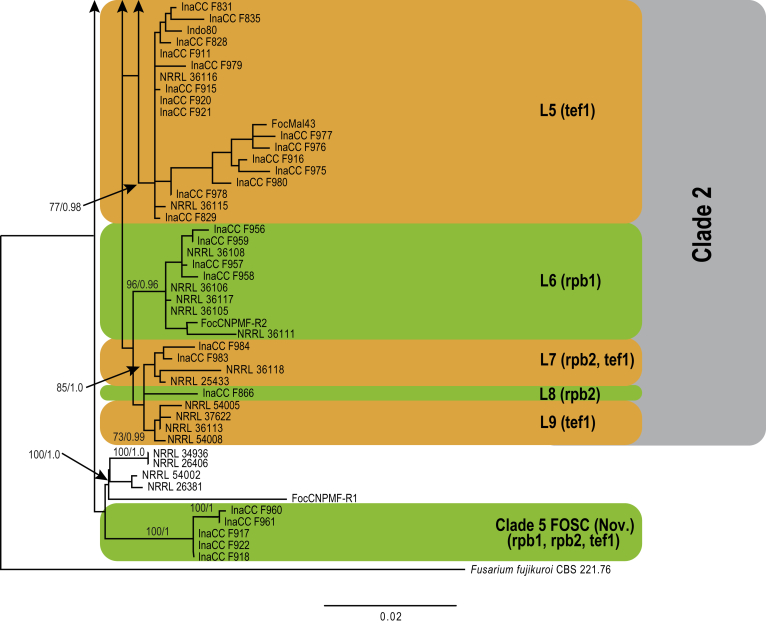
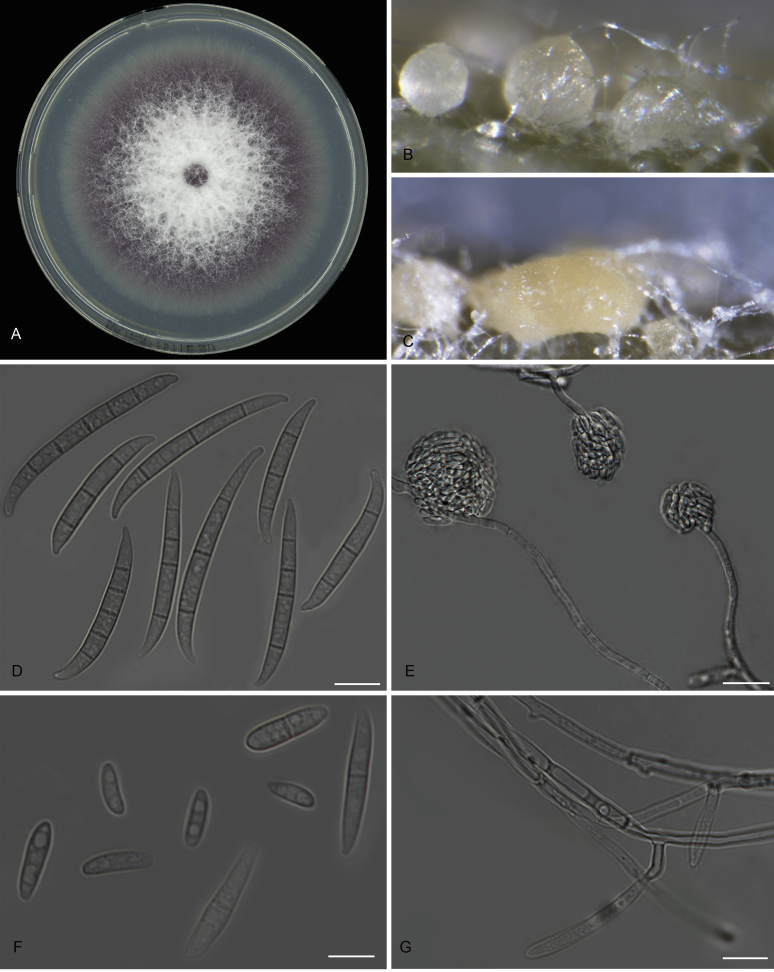
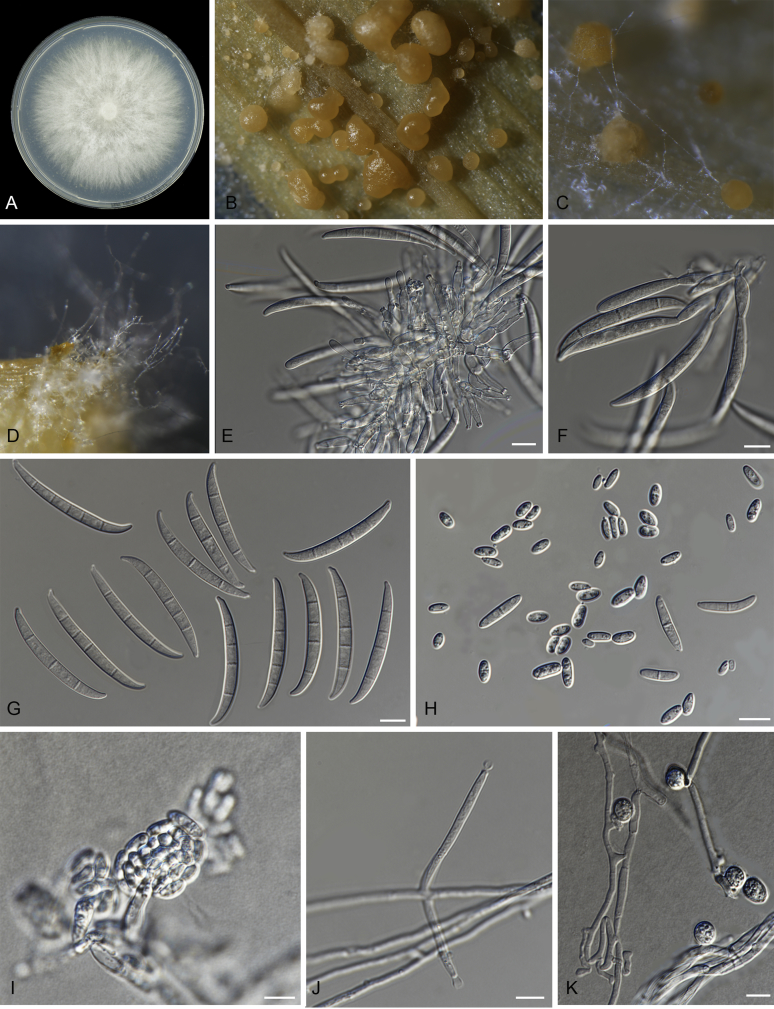
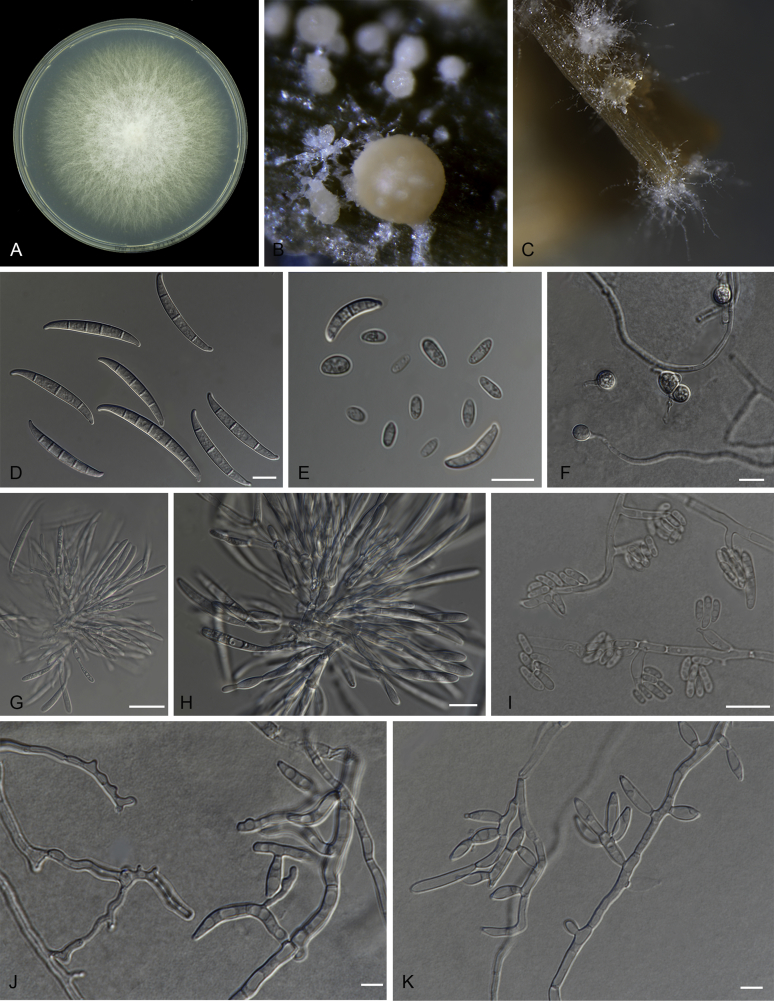
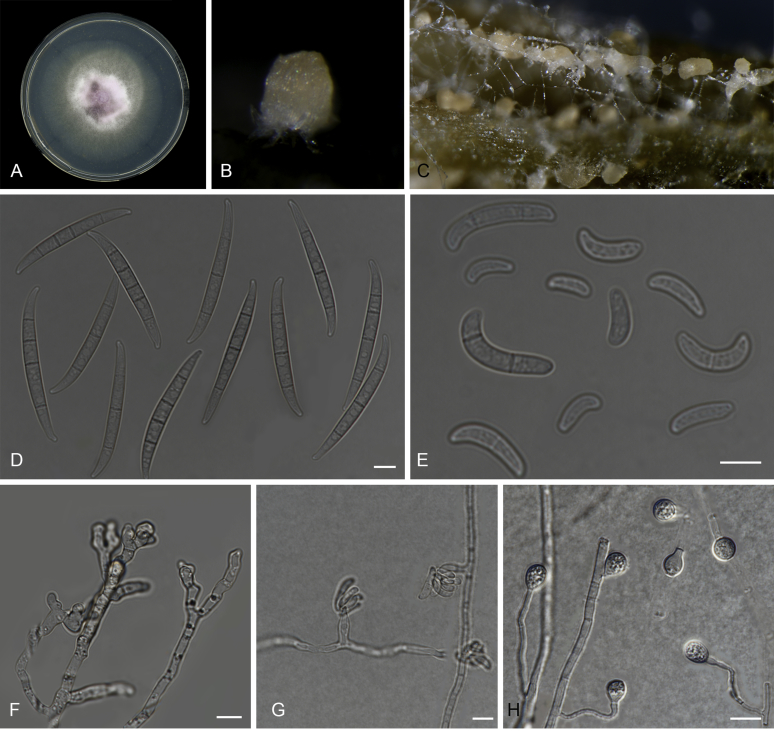
Fusarium oxysporum f. sp. cubense (Foc), the causal agent of Fusarium wilt or Panama disease on banana, is one of the major constraints in banana production worldwide. Indonesia is the centre of origin for wild and cultivated bananas, which likely co-evolved with Foc. This study explored the widest possible genetic diversity of Foc by sampling across Indonesia at 34 geographically and environmentally different locations in 15 provinces at six islands. This resulted in a comprehensive collection of ∼200 isolates from 40 different local banana varieties. Isolates were identified and assessed using sequence analysis of the translation elongation factor-1alpha (tef1), the RNA polymerase II largest subunit (rpb1), and the RNA polymerase II second largest subunit (rpb2). Phylogenetic analyses of these genes allowed the identification of 180 isolates of Fusarium oxysporum f. sp. cubense (Foc), and 20 isolates of the Fusarium fujikuroi species complex (FFSC), the Fusarium incarnatum-equiseti species complex (FIESC), and the Fusarium sambucinum species complex (FSSC). Further analyses, incorporating a worldwide collection of Foc strains, revealed nine independent genetic lineages for Foc, and one novel clade in the Fusarium oxysporum species complex (FOSC). Selected isolates from each lineage were tested on the banana varieties Gros Michel and Cavendish to characterise their pathogenicity profiles. More than 65 % of the isolates were diagnosed as Tropical Race 4 (Foc-TR4) due to their pathogenicity to Cavendish banana, which supports the hypothesis that Foc-TR4 is of Indonesian origin. Nine independent genetic lineages for Foc are formally described in this study. This biodiversity has not been studied since the initial description of Foc in 1919. This study provides a detailed overview of the complexity of Fusarium wilt on banana and its diversity and distribution across Indonesia.
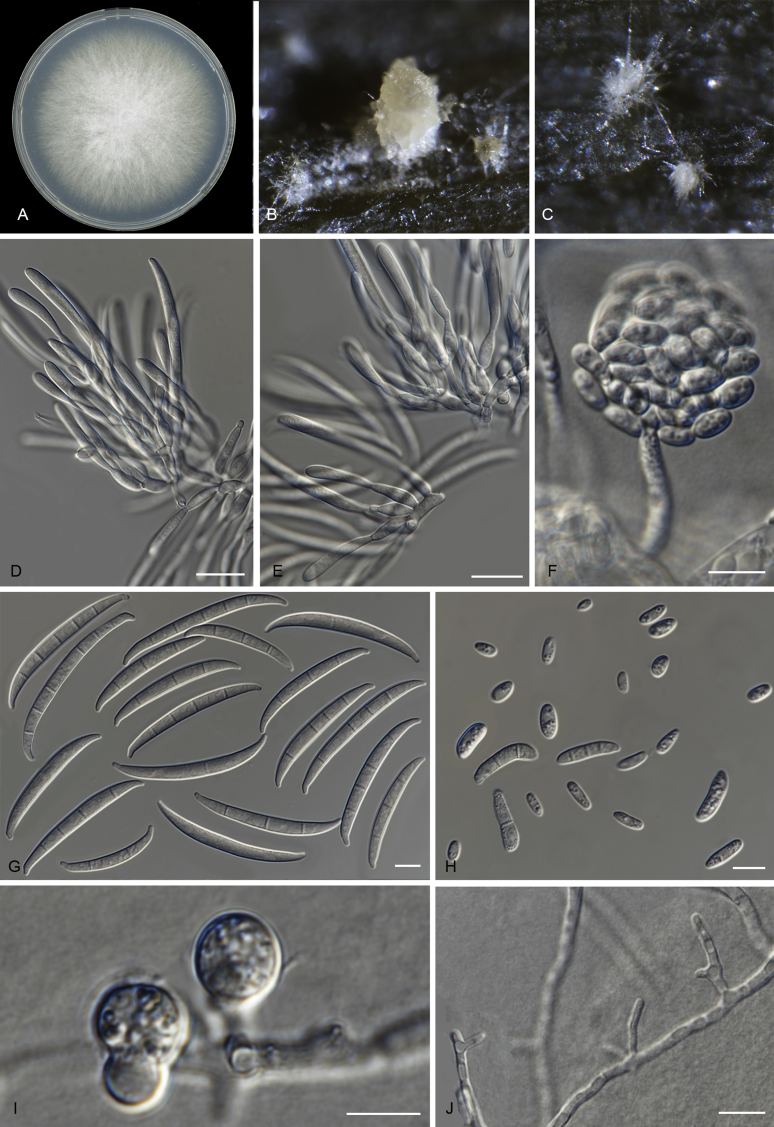
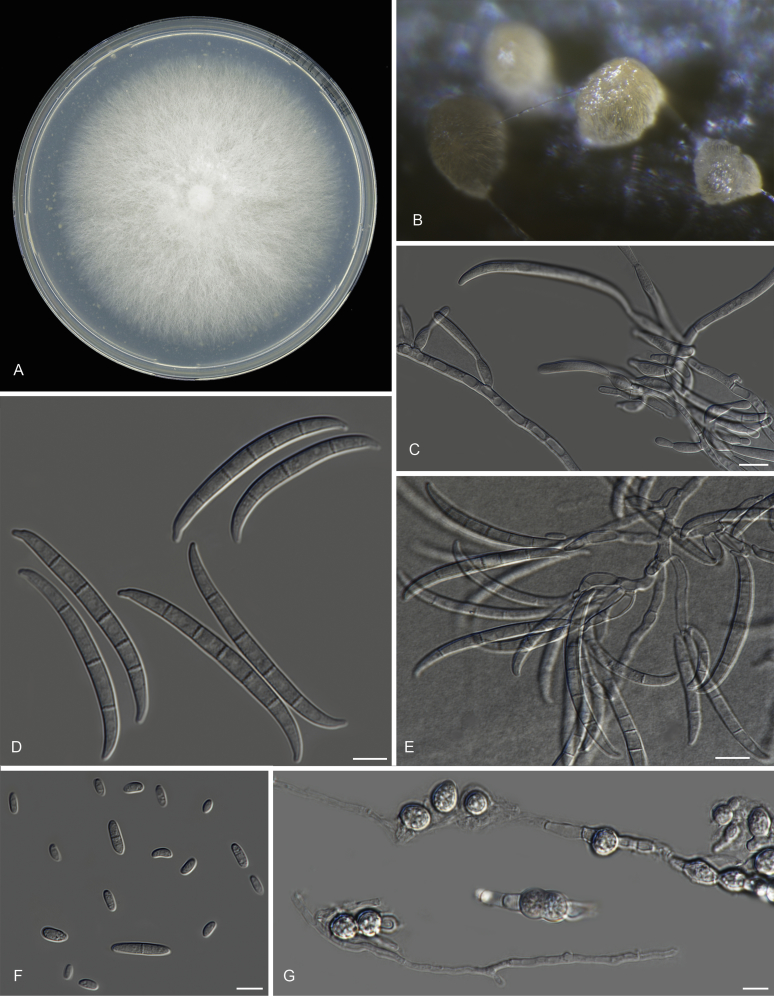
Keywords: 11 New taxa; F. duoseptatum N. Maryani, L. Lombard, Kema & Crous; F. grosmichelii N. Maryani, L. Lombard, Kema & Crous; F. hexaseptatum N. Maryani, L. Lombard, Kema & Crous; F. kalimantanense N. Maryani, L. Lombard, Kema & Crous; F. odoratissimum N. Maryani, L. Lombard, Kema & Crous; F. phialophorum N. Maryani, L. Lombard, Kema & Crous; F. purpurascens N. Maryani, L. Lombard, Kema & Crous; F. sangayamense N. Maryani, L. Lombard, Kema & Crous; F. tardichlamydosporum N. Maryani, L. Lombard, Kema & Crous; F. tardicrescens N. Maryani, L. Lombard, Kema & Crous; Fusarium cugenangense N. Maryani, L. Lombard, Kema & Crous; Morphology; New species; Panama disease; Pathogenicity; Tropical Race 4.
Figures





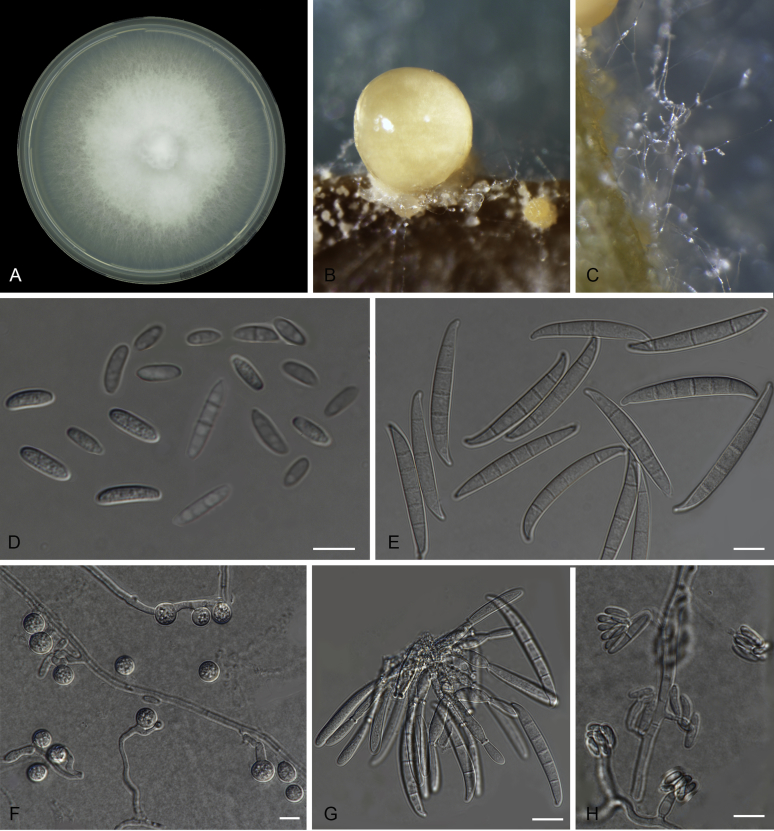
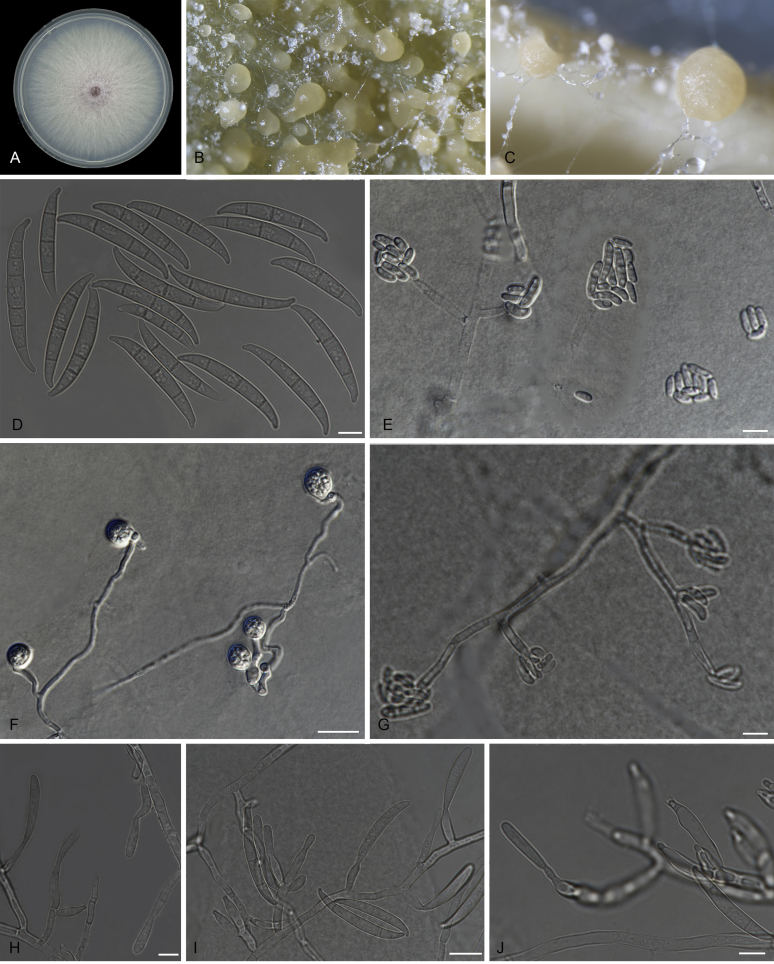
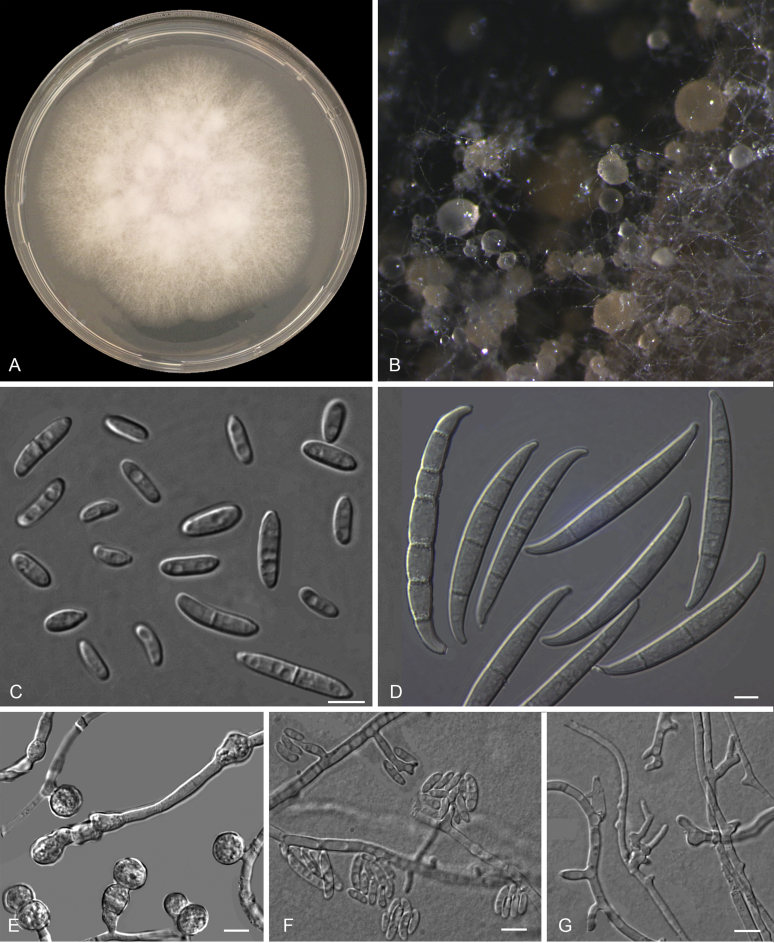
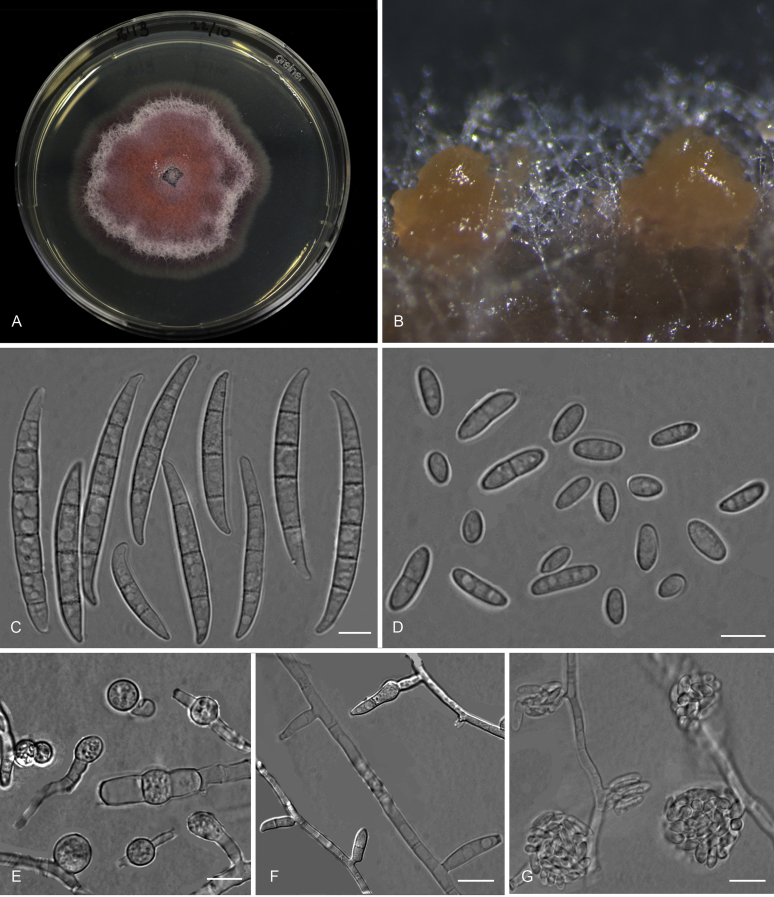
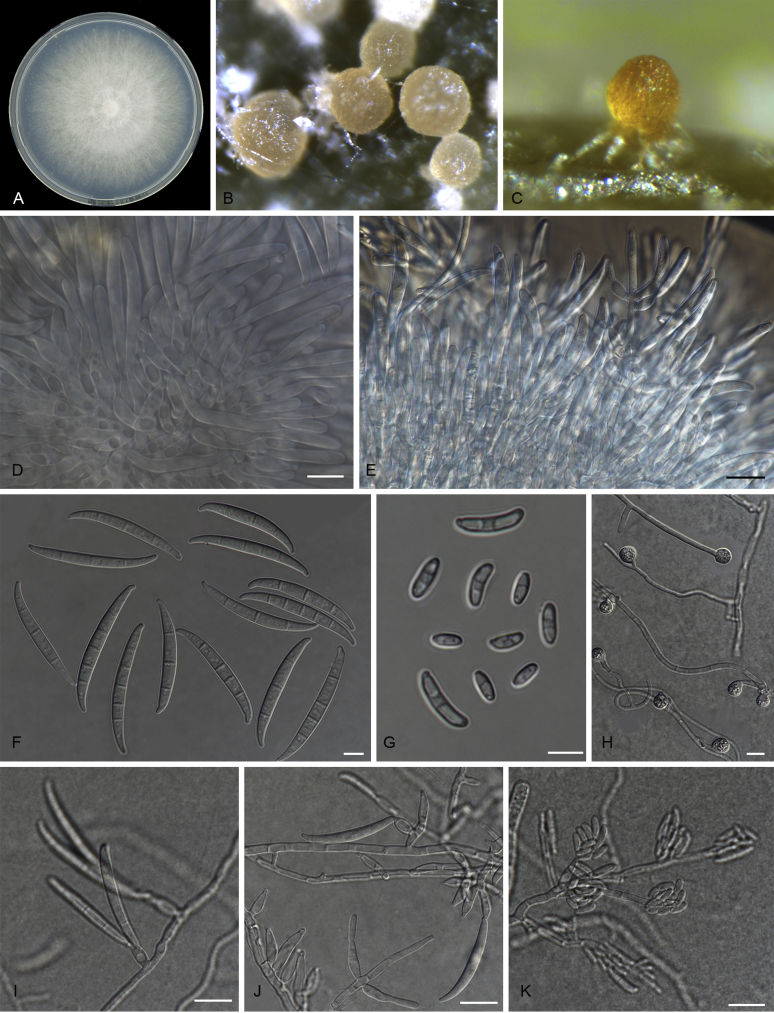
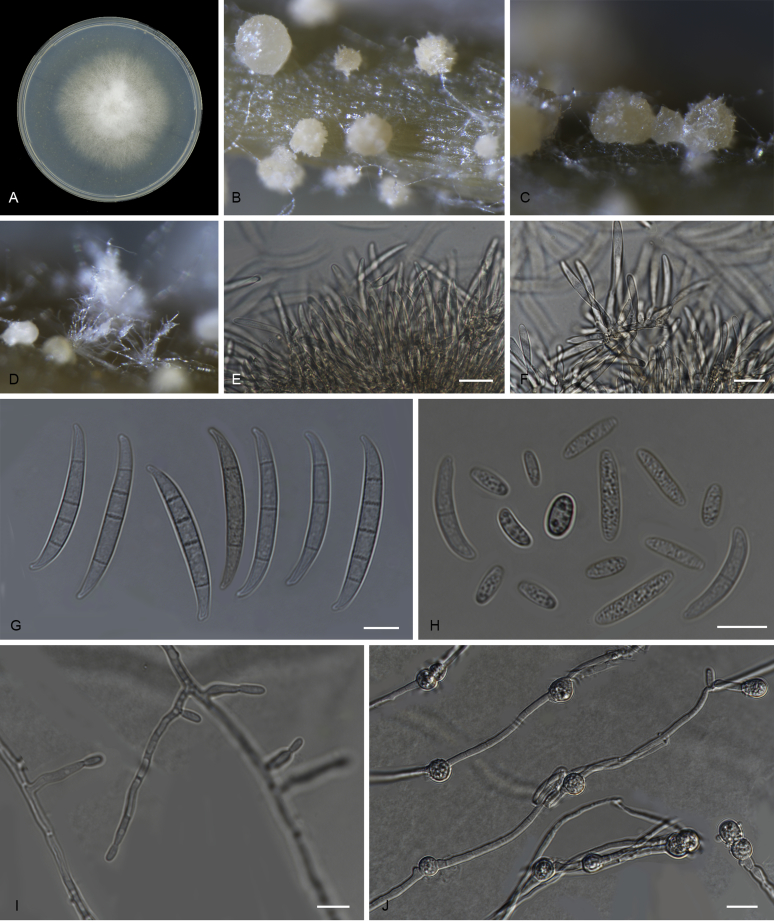

References
-
- Baayen R.P., O'Donnell K., Bonants P.J.M. Gene genealogies and AFLP analyses in the Fusarium oxysporum complex identify monophyletic and nonmonophyletic formae speciales causing wilt and rot disease. Phytopathology. 2000;90:891–900. - PubMed
-
- Banke S., Peschon A., McDonald B.A. Phylogenetic analysis of globally distributed Mycosphaerella graminicola populations based on three DNA sequence loci. Fungal Genetics and Biology. 2004;41:226–238. - PubMed
-
- Bentley S., Pegg K.G., Dale J.L. Genetic variation among a world-wide collection of isolates of Fusarium oxysporum f. sp. cubense analysed by RAPD-PCR fingerprinting. Mycological Research. 1995;99:1378–1384.
-
- Bentley S., Pegg K.G., Moore N.Y. Genetic variation among vegetative compatibility groups of Fusarium oxysporum f. sp. cubense analyzed by DNA fingerprinting. Phytopathology. 1998;88:1283–1293. - PubMed
-
- Boehm E., Ploetz R., Kistler C.H. Statistical analysis of electrophoretic karyotype variation among vegetative compatibility groups of Fusarium oxysporum f. sp. cubense. MPMI-Molecular Plant Microbe Interactions. 1994;7:196–207.
LinkOut - more resources
Full Text Sources
Other Literature Sources