

The suppressive cap-binding complex factor 4EIP is required for normal differentiation
- PMID: 30124912
- PMCID: PMC6158607
- DOI: 10.1093/nar/gky733
The suppressive cap-binding complex factor 4EIP is required for normal differentiation
Abstract
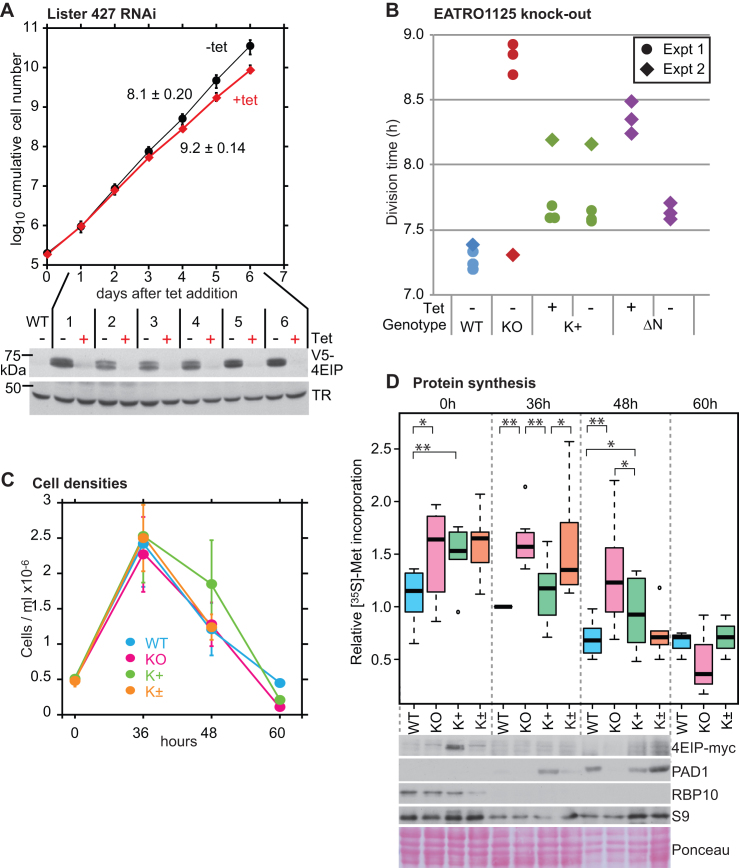
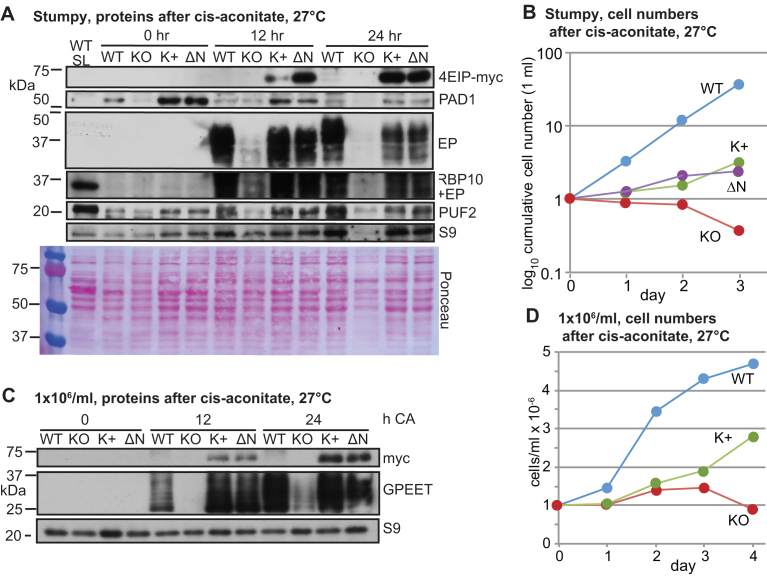
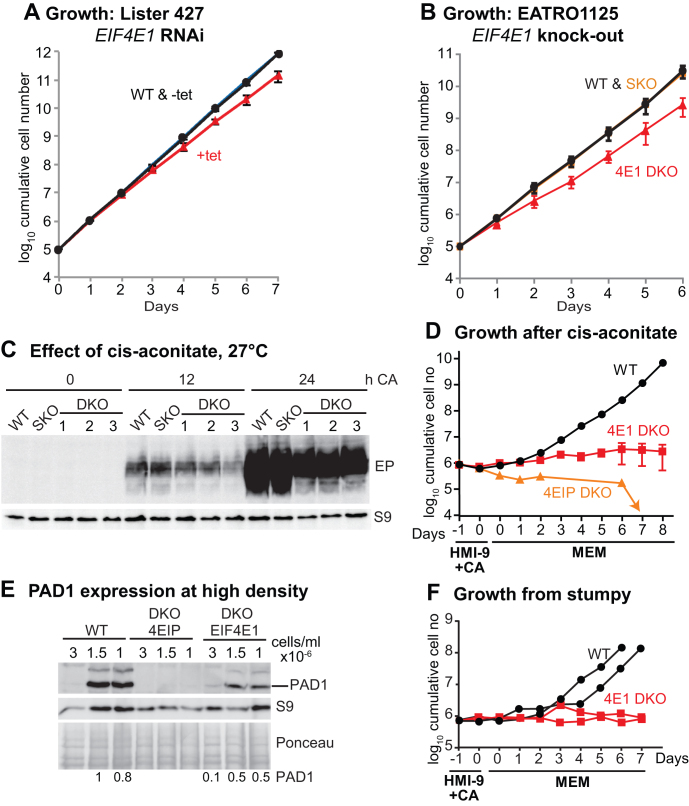
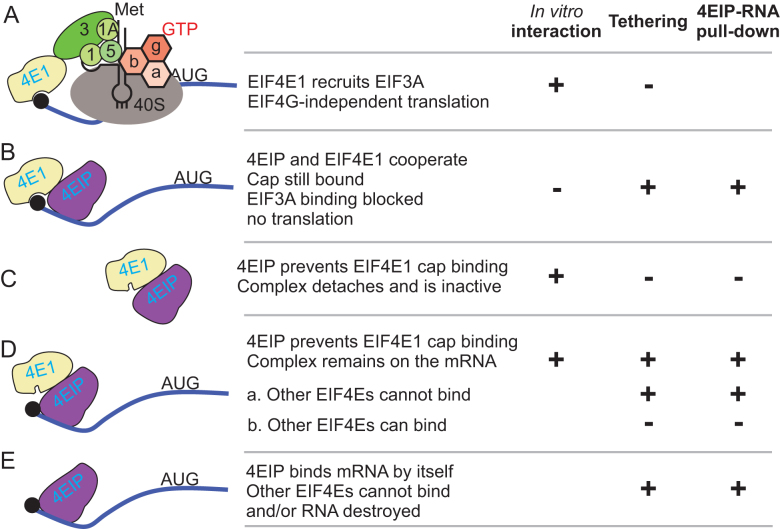
Trypanosoma brucei live in mammals as bloodstream forms and in the Tsetse midgut as procyclic forms. Differentiation from one form to the other proceeds via a growth-arrested stumpy form with low messenger RNA (mRNA) content and translation. The parasites have six eIF4Es and five eIF4Gs. EIF4E1 pairs with the mRNA-binding protein 4EIP but not with any EIF4G. EIF4E1 and 4EIP each inhibit expression when tethered to a reporter mRNA, but while tethered EIF4E1 suppresses only when 4EIP is present, suppression by tethered 4EIP does not require the interaction with EIF4E1. In growing bloodstream forms, 4EIP is preferentially associated with unstable mRNAs. Bloodstream- or procyclic-form trypanosomes lacking 4EIP have only a marginal growth disadvantage. Bloodstream forms without 4EIP are, however, defective in translation suppression during stumpy-form differentiation and cannot subsequently convert to growing procyclic forms. Intriguingly, the differentiation defect can be complemented by a truncated 4EIP that does not interact with EIF4E1. In contrast, bloodstream forms lacking EIF4E1 have a growth defect, stumpy formation seems normal, but they appear unable to grow as procyclic forms. We suggest that 4EIP and EIF4E1 fine-tune mRNA levels in growing cells, and that 4EIP contributes to translation suppression during differentiation to the stumpy form.
Figures

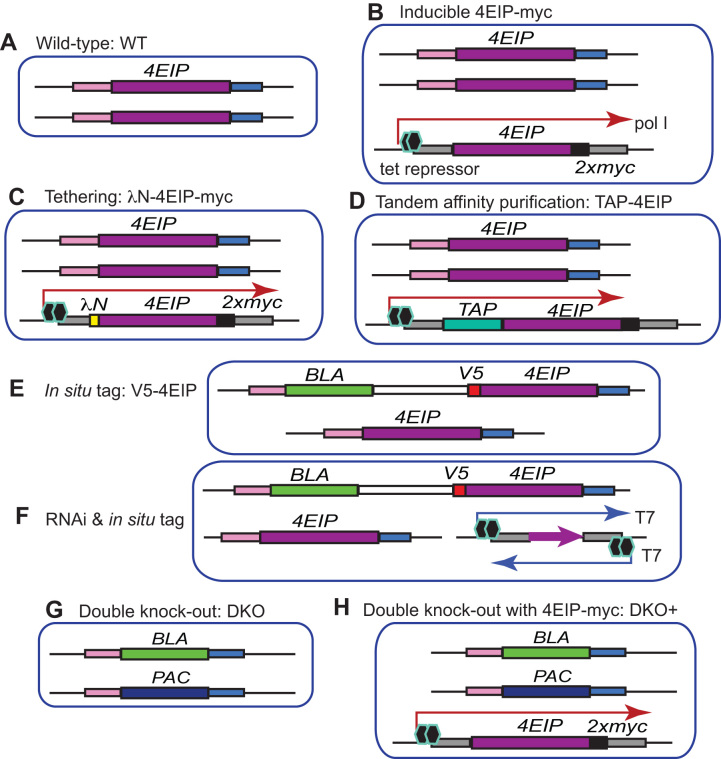
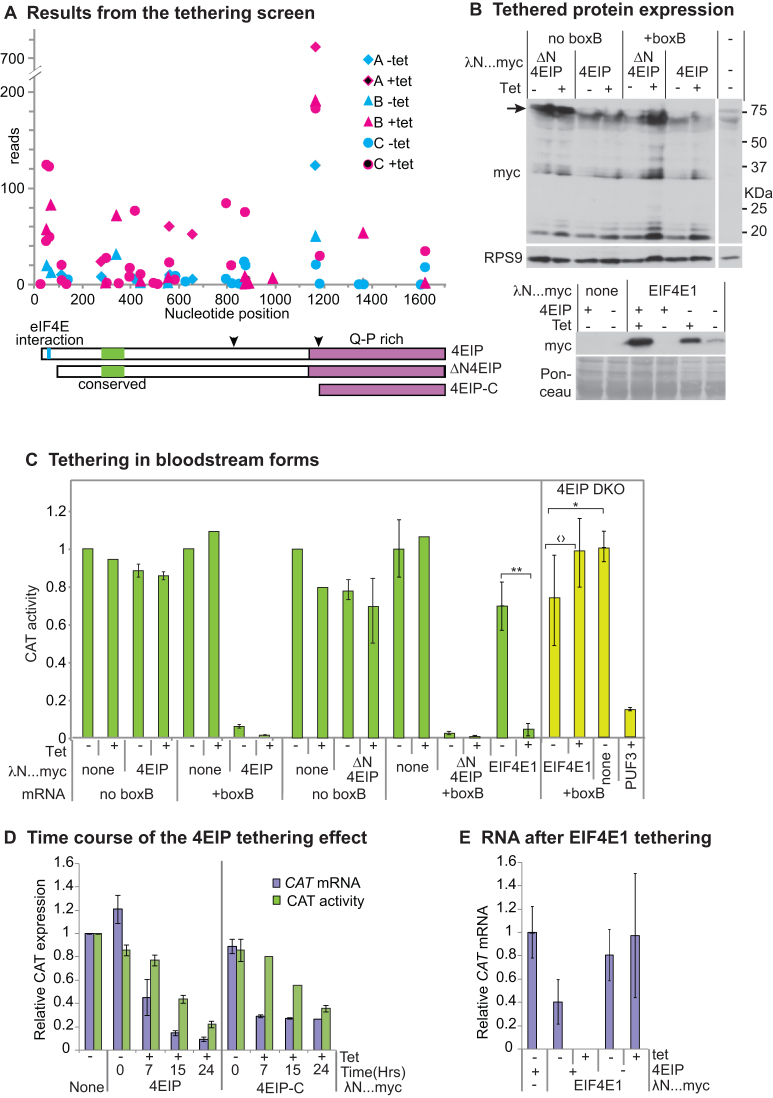
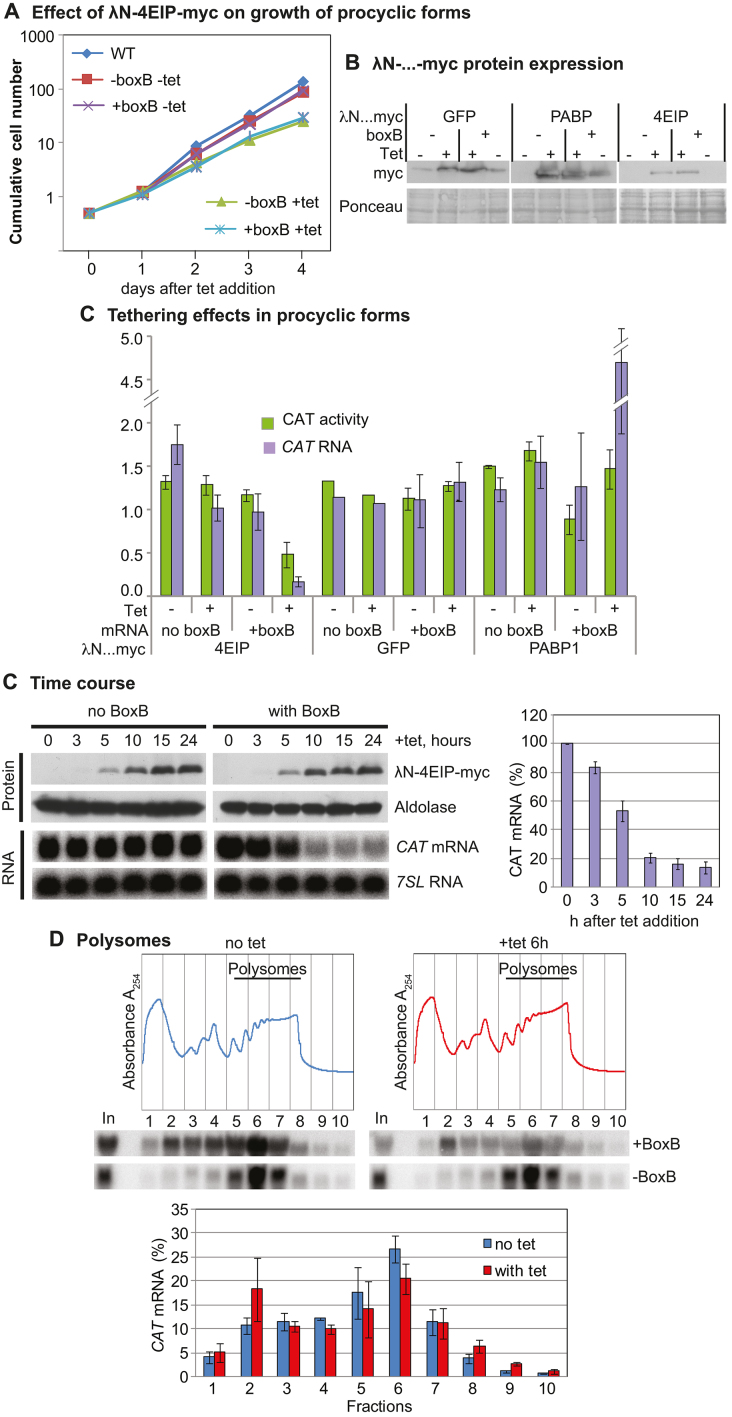
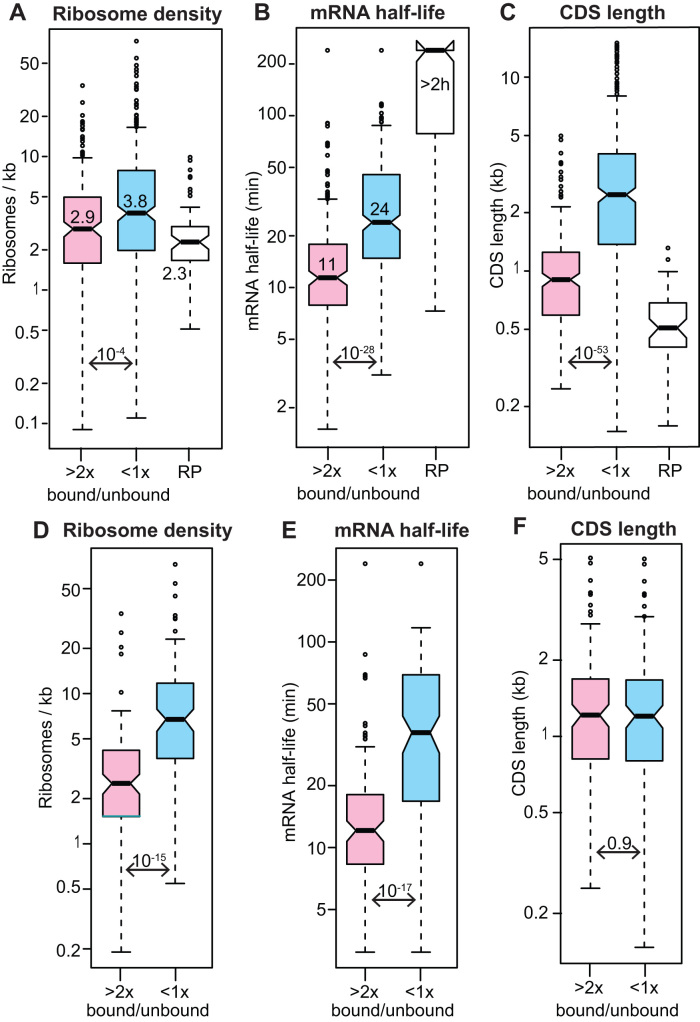
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
