

Environmental triggers for photosynthetic protein turnover determine the optimal nitrogen distribution and partitioning in the canopy
- PMID: 30124935
- PMCID: PMC6519421
- DOI: 10.1093/jxb/ery308
Environmental triggers for photosynthetic protein turnover determine the optimal nitrogen distribution and partitioning in the canopy
Abstract
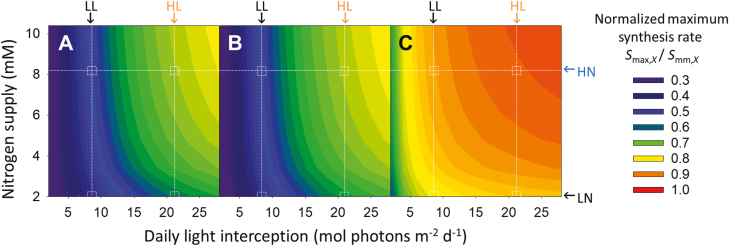
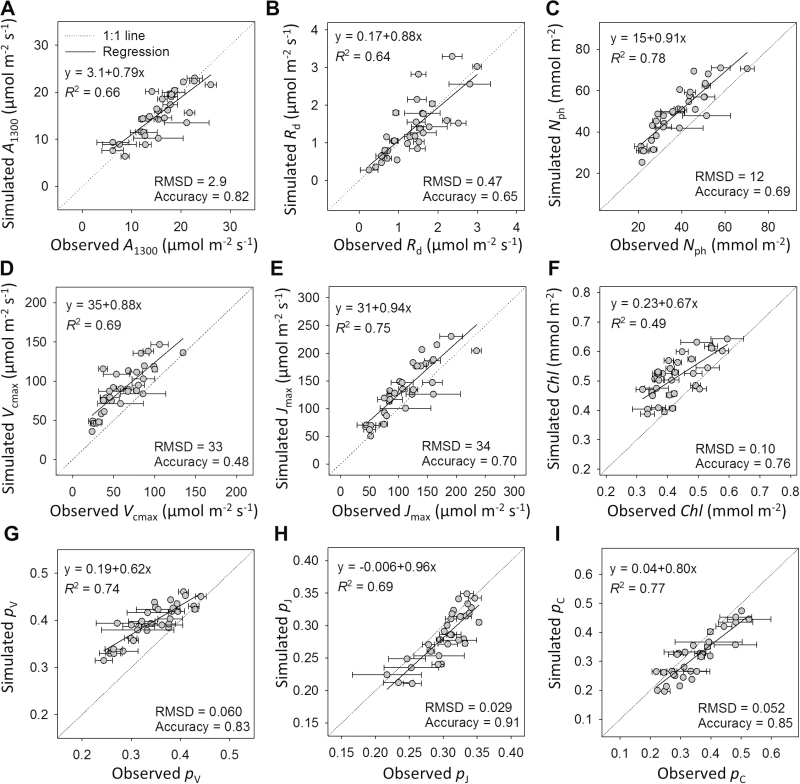
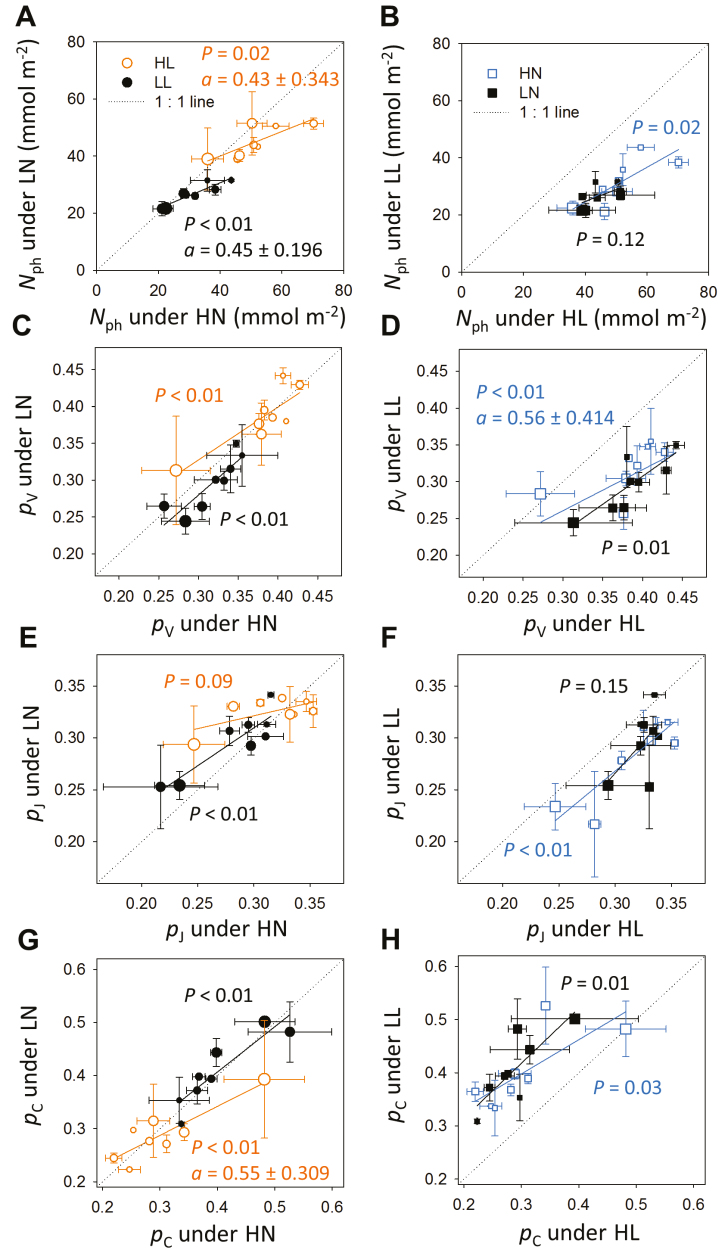
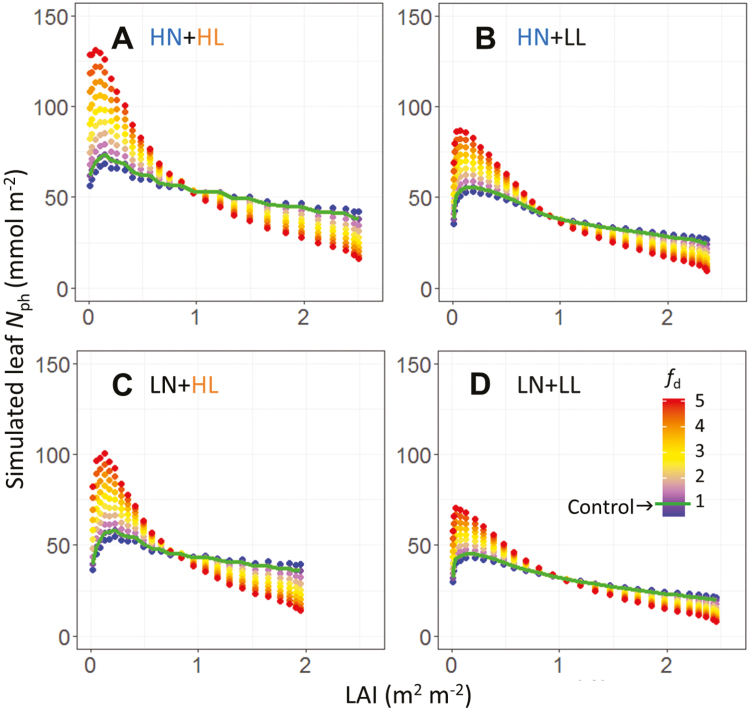
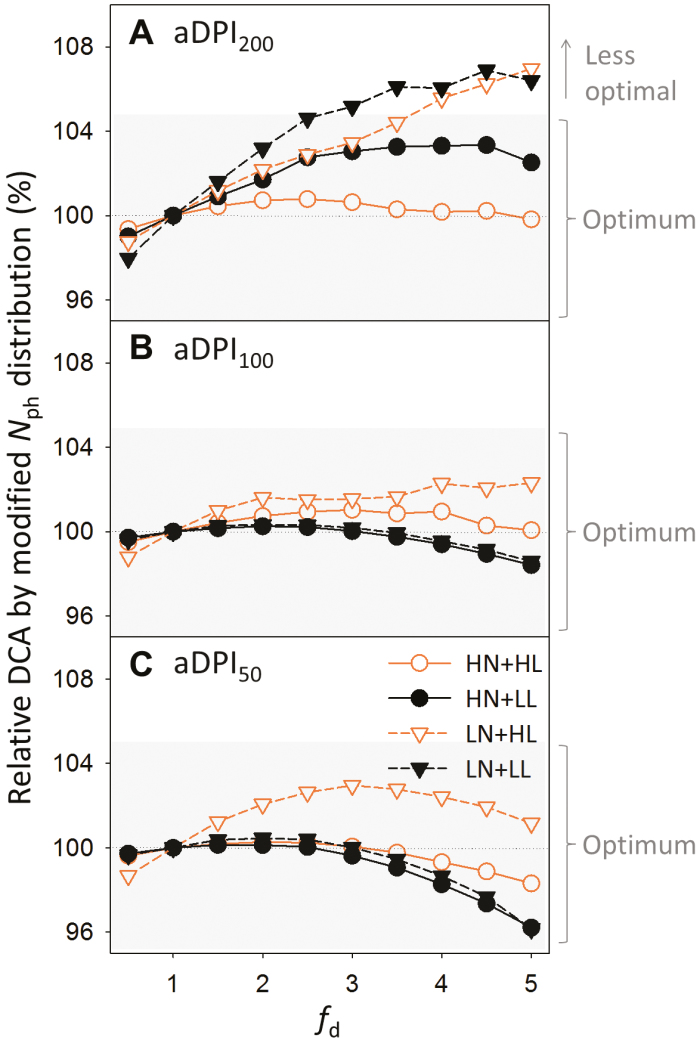
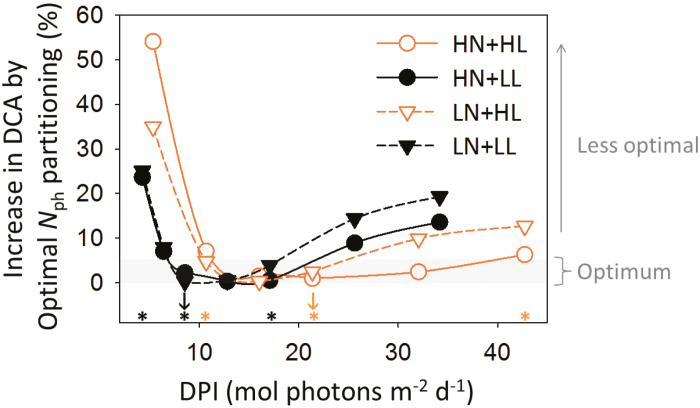
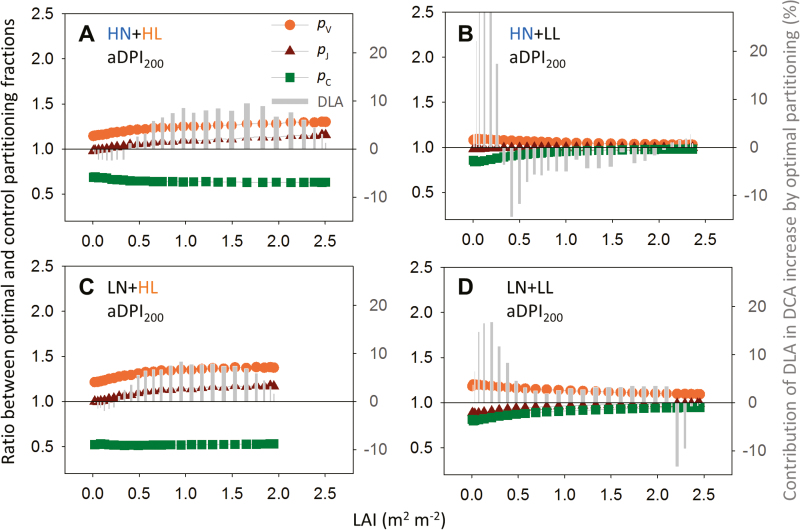
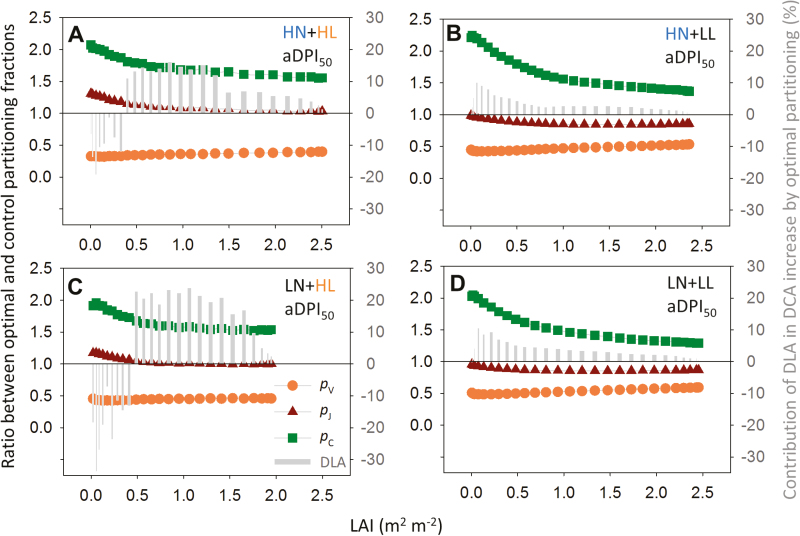
Plants continually adjust the photosynthetic functions in their leaves to fluctuating light, thereby optimizing the use of photosynthetic nitrogen (Nph) at the canopy level. To investigate the complex interplay between external signals during the acclimation processes, a mechanistic model based on the concept of protein turnover (synthesis and degradation) was proposed and parameterized using cucumber grown under nine combinations of nitrogen and light in growth chambers. Integrating this dynamic model into a multi-layer canopy model provided accurate predictions of photosynthetic acclimation of greenhouse cucumber canopies grown under high and low nitrogen supply in combination with day-to-day fluctuations in light at two different levels. This allowed us to quantify the degree of optimality in canopy nitrogen use for maximizing canopy carbon assimilation, which was influenced by Nph distribution along canopy depth or Nph partitioning between functional pools. Our analyses suggest that Nph distribution is close to optimum and Nph reallocation is more important under low nitrogen. Nph partitioning is only optimal under a light level similar to the average light intensity during acclimation, meaning that day-to-day light fluctuations inevitably result in suboptimal Nph partitioning. Our results provide insights into photoacclimation and can be applied to crop model improvement.
Keywords: Functional partitioning; light; mechanistic model; nitrogen reallocation; nitrogen supply; optimal; photosynthetic acclimation.
© The Author(s) 2018. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Anten NP , Miyazawa K , Hikosaka K , Nagashima H , Hirose T. 1998. Leaf nitrogen distribution in relation to leaf age and photon flux density in dominant and subordinate plants in dense stands of a dicotyledonous herb. Oecologia 113, 314–324. - PubMed
-
- Anten NP , Schieving F , Werger MJ. 1995. Patterns of light and nitrogen distribution in relation to whole canopy carbon gain in C3 and C4 mono- and dicotyledonous species. Oecologia 101, 504–513. - PubMed
-
- Bathellier C , Tcherkez G , Lorimer GH , Farquhar GD. 2018. Rubisco is not really so bad. Plant, Cell & Environment 41, 705–716. - PubMed
-
- Boote KJ , Jones JW , White JW , Asseng S , Lizaso JI. 2013. Putting mechanisms into crop production models. Plant, Cell & Environment 36, 1658–1672. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
