

doi: 10.1093/nar/gky735.
i-Motif DNA: structural features and significance to cell biology
Affiliations
- PMID: 30124962
- PMCID: PMC6144788
- DOI: 10.1093/nar/gky735
Item in Clipboard
i-Motif DNA: structural features and significance to cell biology
Nucleic Acids Res.
.
Abstract
The i-motif represents a paradigmatic example of the wide structural versatility of nucleic acids. In remarkable contrast to duplex DNA, i-motifs are four-stranded DNA structures held together by hemi- protonated and intercalated cytosine base pairs (C:C+). First observed 25 years ago, and considered by many as a mere structural oddity, interest in and discussion on the biological role of i-motifs have grown dramatically in recent years. In this review we focus on structural aspects of i-motif formation, the factors leading to its stabilization and recent studies describing the possible role of i-motifs in fundamental biological processes.
Figures
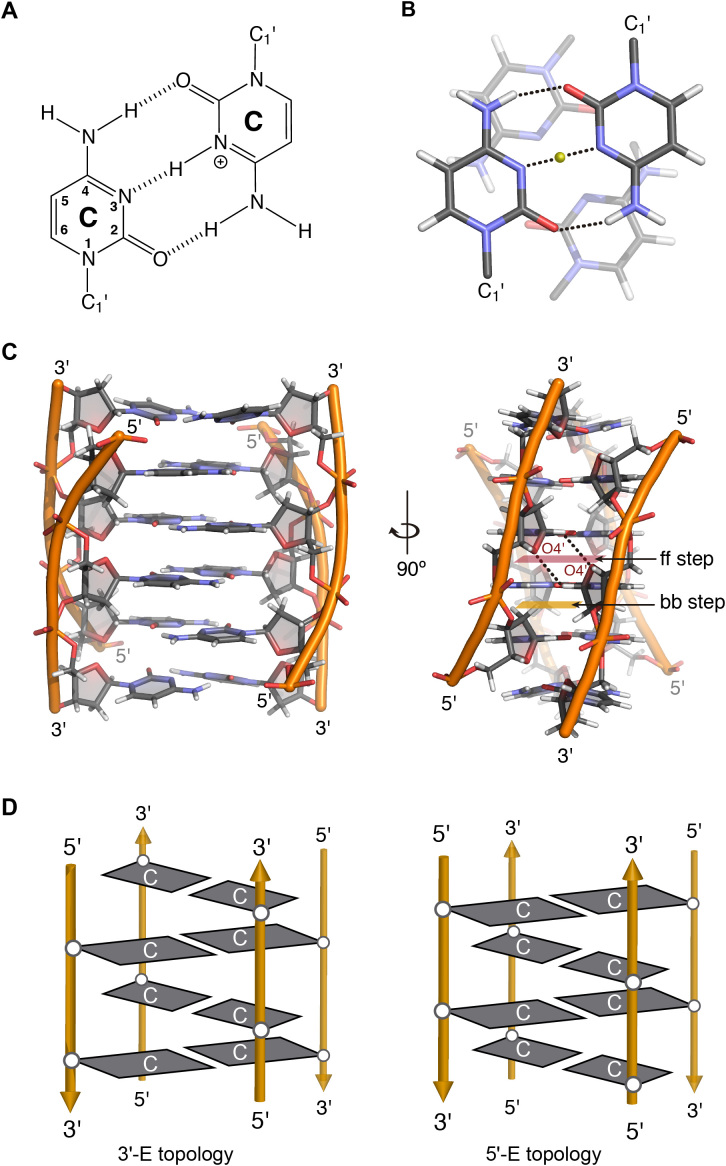
(A) C:C+ base pair. (B) Stacking between intercalating C:C+ base pairs. (C) Two views of the 3D structure of a tetramolecular i-motif (PDB 1YBR), face-to-face and back-to-back steps are highlighted and two hydrogen bonds between O4′ and H1′ are marked with black dashed lines. (D) Scheme representing the 3′E (left) and 5′E (right) intercalation topologies of an intermolecular i-motif structure.
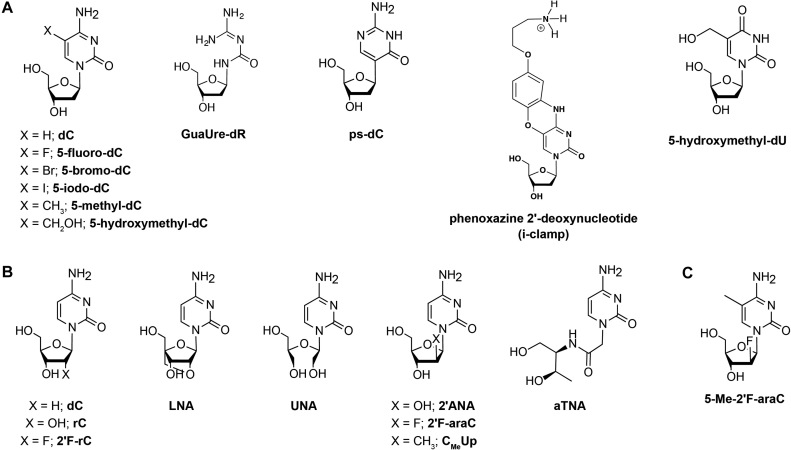
Nucleoside modifications introduced in i-motif structures. (A) Nucleobase modifications. (B) Sugar modifications and (C) A nucleoside analogue possessing both sugar and nucleobase modifications. d: deoxyribose, r: ribose and ara: arabinose sugar.
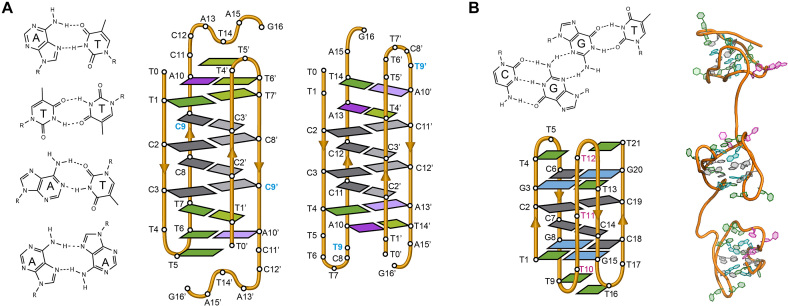
(A) Several base pairs other than C:C+ that have been observed in i-motif structures (from top to bottom; A:T Hoogsteen, T:T, A:T reverse Watson-Crick and A:A) (Left). Schematic view of the dimeric i-motif structures formed by the two variants of centromeric A-box sequence (63) showing the presence of T:T and A:T reverse Watson–Crick and Hoogsteen base pairs flanking the set of intercalated C:C+ base pairs (Right). The residue in blue denotes the only difference in the primary structure of the two sequences. (B) G:C:G:T minor groove tetrad (Top). Schematic representation of the minimal i-motif structure stabilized by two C:C+ base pairs and two flanking G:C:G:T minor groove tetrads (Bottom) and representation of these structures in tandem (Right) (53).
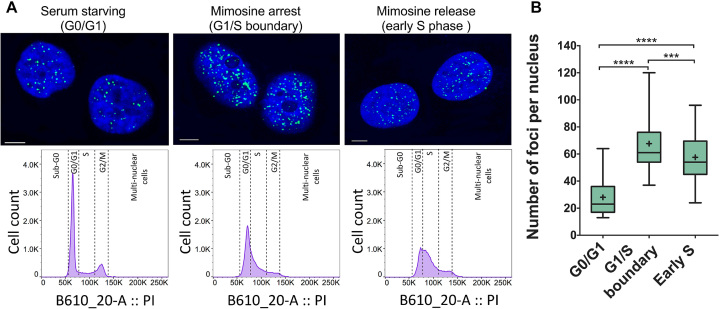
(A) Confocal microscopy images showing fluorescent foci of i-motif recognized by iMab antibody in the nuclei of HeLa cells arrested at different stages of cell cycle. Flow cytometry diagrams show the population of synchronized and propidium iodide stained cells in each stage. (B) Box plot graph showing the quantification of iMab foci in each phase. Boxes represent 25th to 75th percentile. Horizontal line and ‘+’ symbol inside the box represent medians and means, respectively. Whiskers indicate lower and highest values registered in each case. ***P < 0.001, ****P < 0.0001. Reprinted with permission from Zeraati, M., Langley, D.B., Schofield, P., Moye, A.L., Rouet, R., Hughes, W.E., Bryan, T.M., Dinger, M.E. and Christ, D. (2018) I-motif DNA structures are formed in the nuclei of human cells. Nature chemistry, 10, 631–637. Copyright (2018) Springer Nature.
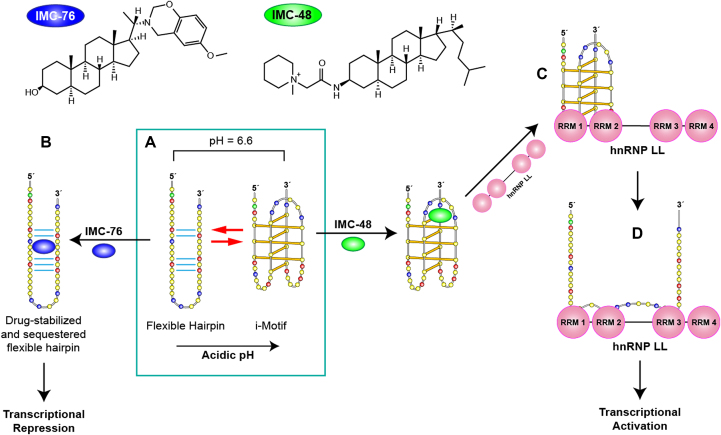
(A) Structures of IMC-48 and IMC-76 ligands, specific for BCL2 i-motif and BCL2 hairpin, respectively. (B) BCL2 promoter sequence can adopt two different major conformations which are in a pH-dependent equilibrium. Acidic conditions lead to formation of an i-motif whereas at pH 6.6 there is a mixture of hairpin and i-motif conformations. IMC-76 binds to the hairpin structure shifting the equilibrium towards the hairpin structure which leads to transcriptional repression (A to B) whereas IMC-48 binds to the central loop of the BCL2 i-motif stabilizing the structure that upon binding of hnRNP LL via RRMs 1 and 2 of the protein (A to C) promotes the unfolding of the i-motif and the activation of BCL2 transcription (C to D). Adapted with permission from Kang, H.J., Kendrick, S., Hecht, S.M. and Hurley, L.H. (2014) The Transcriptional Complex Between the BCL2 i-Motif and hnRNP LL Is a Molecular Switch for Control of Gene Expression That Can Be Modulated by Small Molecules. J Am Chem Soc, 136, 4172–4185. Copyright (2014) American Chemical Society.
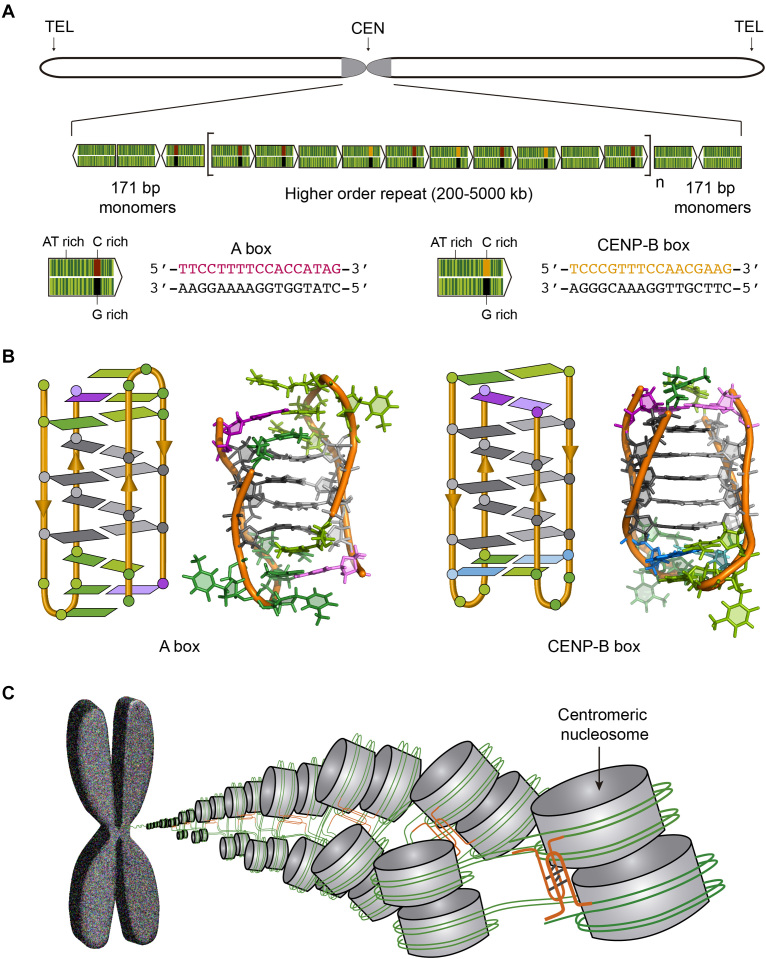
(A) Human chromosome showing the centromere in grey and the structure of the human centromeric alpha satellite DNA. The 171 bp monomers (green arrows) are AT-rich sequences that can contain either A box (red and black) or CENP-B box (orange and black). The monomers are tandemly repeated and form a higher order repeat, which in turn appears repeated along the centromere. (B) Schematic and cartoon structures of the dimeric i-motifs formed by truncated A box (PDB ID: 2MRZ) and CENP-B box (PDB ID: 1C11). Cytosines are represented in grey, thymines in green, adenines in magenta and guanines in blue. (C) Proposed model for nucleosome organization at the centromere. The i-motifs formed by C-rich sequences of A and CENP-B boxes (orange) would maintain the structural organization of the centromere.
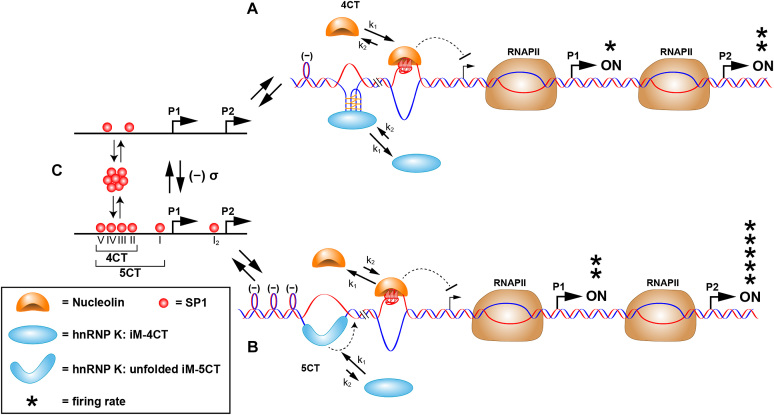
Proposed scheme for the molecular mechanosensor mechanism for differential control of MYC expression through the NHE III1. (A) Low SP1 occupancy of duplex promoter binding sites (as shown in (C), upper) results in low negative supercoiling and basal levels of transcription from the P1 and P2 promoters. In this scenario, hnRNP K can only access the central and lateral loops in the i-motif that bind to two of the KH domains; as such only the 4CT element is accessed, and nucleolin binding predominates over hnRNP K binding to the equilibrating G-quadruplex and i-motif, which are mutually exclusive. (B) At higher occupancy levels of SP1 to the duplex promoter binding sites (as shown in (C), lower), which results in enhanced negative supercoiling, the 5CT element is now fully melted, and hnRNP K forms a thermodynamically stable complex binding through the addition of the CT element. The binding affinity of hnRNP K to the unfolded i-motif and the additional CT element involving three KH domains now exceeds that of nucleolin to the G-quadruplex, and MYC expression from P1 and P2 is at peak levels. Reprinted with permission from Sutherland, C., Cui, Y.X., Mao, H.B. and Hurley, L.H. (2016) A Mechanosensor Mechanism Controls the G-Quadruplex/i-Motif Molecular Switch in the MYC Promoter NHE III1. J Am Chem Soc, 138, 14138–14151. Copyright (2016) American Chemical Society.
References
-
- Watson J.D., Crick F.H.C.. Molecular structure of nucleic acids - a structure for deoxyribose nucleic acid. Nature. 1953; 171:737–738. - PubMed
-
- Du Y., Zhou X.. Targeting non-B-form DNA in living cells. Chem. Rec. 2013; 13:371–384. - PubMed
-
- Choi J., Majima T.. Conformational changes of non-B DNA. Chem. Soc. Rev. 2011; 40:5893–5909. - PubMed
-
- Sen D., Gilbert W.. Formation of parallel 4-Stranded complexes by Guanine-Rich motifs in DNA and its implications for meiosis. Nature. 1988; 334:364–366. - PubMed
-
- Gehring K., Leroy J.L., Gueron M.. A tetrameric DNA-structure with protonated cytosine.cytosine Base-Pairs. Nature. 1993; 363:561–565. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
