

Genetic Code Expansion in Animals
- PMID: 30125487
- PMCID: PMC7026698
- DOI: 10.1021/acschembio.8b00520
Genetic Code Expansion in Animals
Abstract
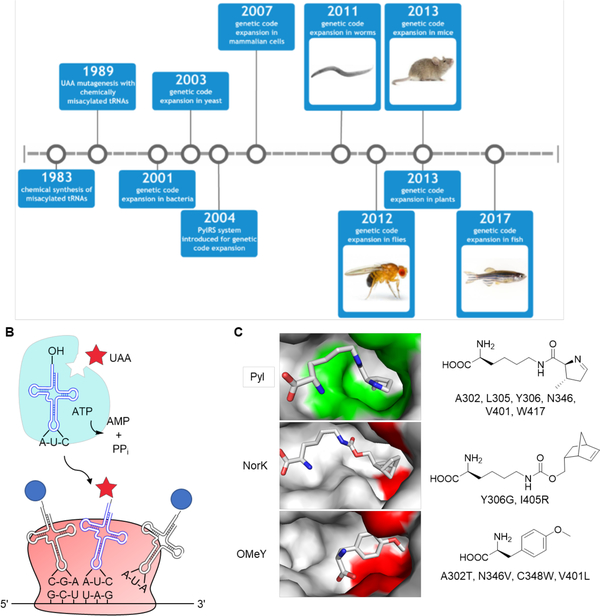
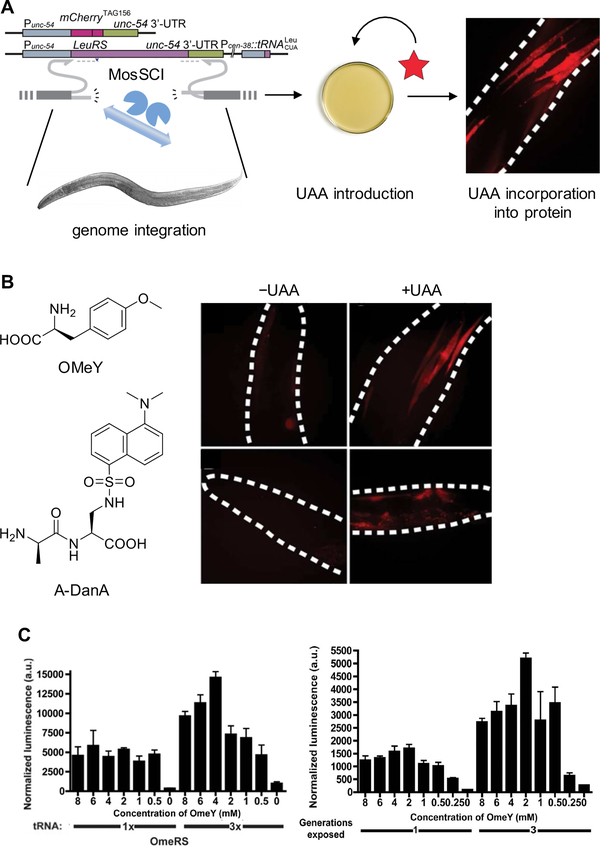
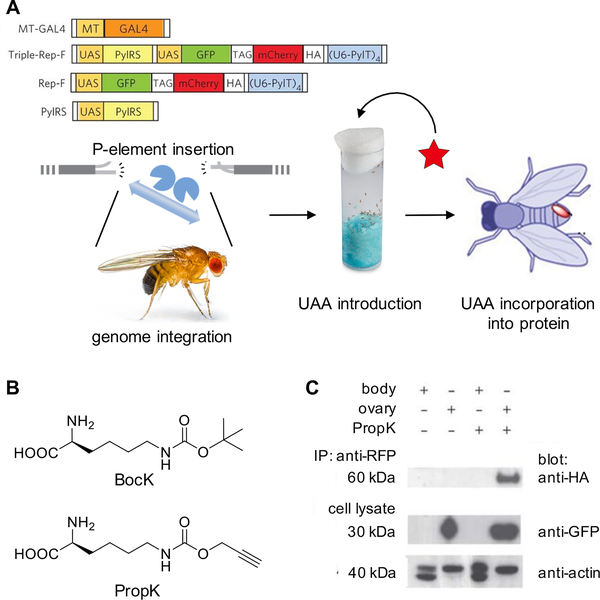
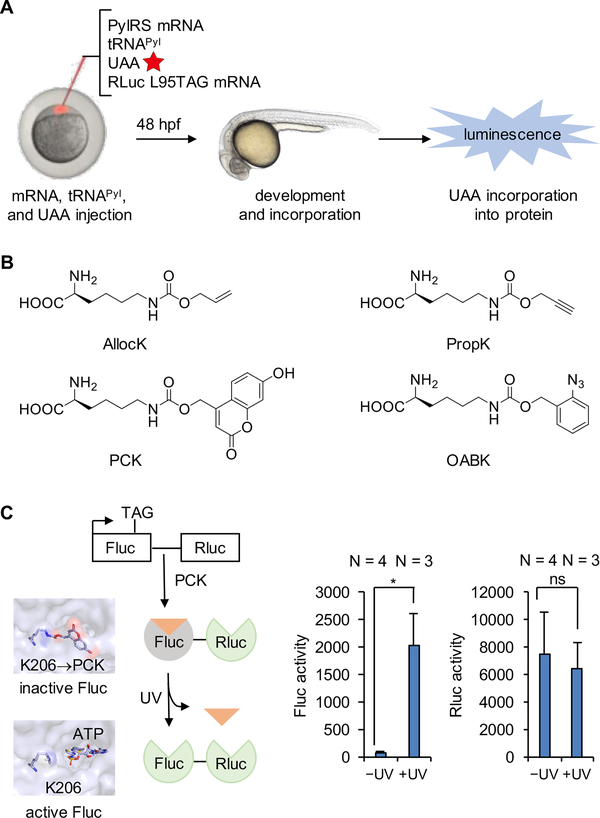
Expanding the genetic code to enable the incorporation of unnatural amino acids into proteins in biological systems provides a powerful tool for studying protein structure and function. While this technology has been mostly developed and applied in bacterial and mammalian cells, it recently expanded into animals, including worms, fruit flies, zebrafish, and mice. In this review, we highlight recent advances toward the methodology development of genetic code expansion in animal model organisms. We further illustrate the applications, including proteomic labeling in fruit flies and mice and optical control of protein function in mice and zebrafish. We summarize the challenges of unnatural amino acid mutagenesis in animals and the promising directions toward broad application of this emerging technology.
Conflict of interest statement
Competing Interests
The authors declare no competing financial interests.
Figures
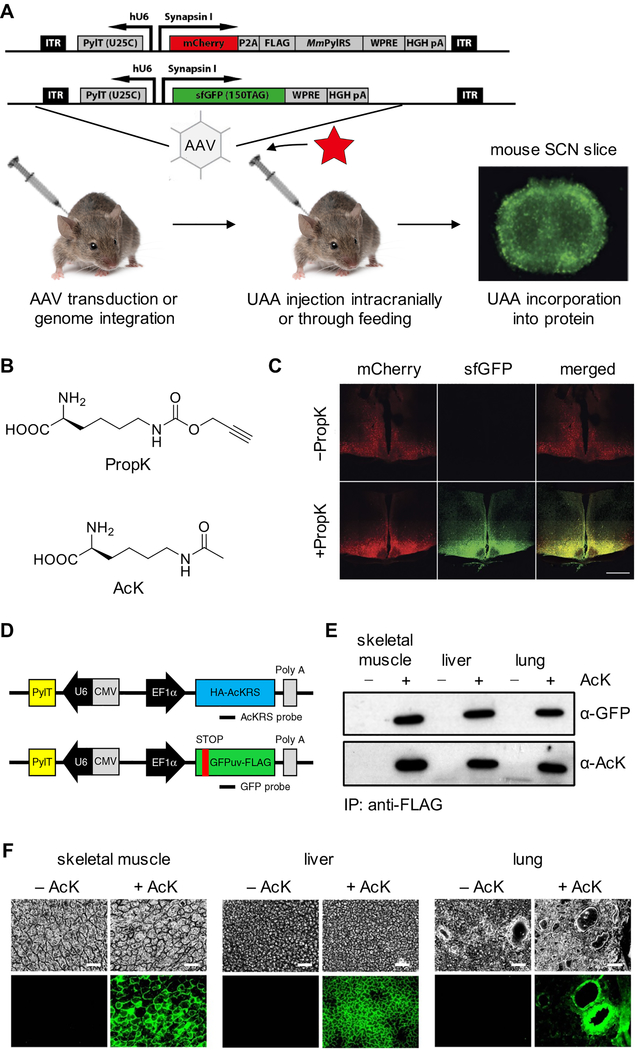
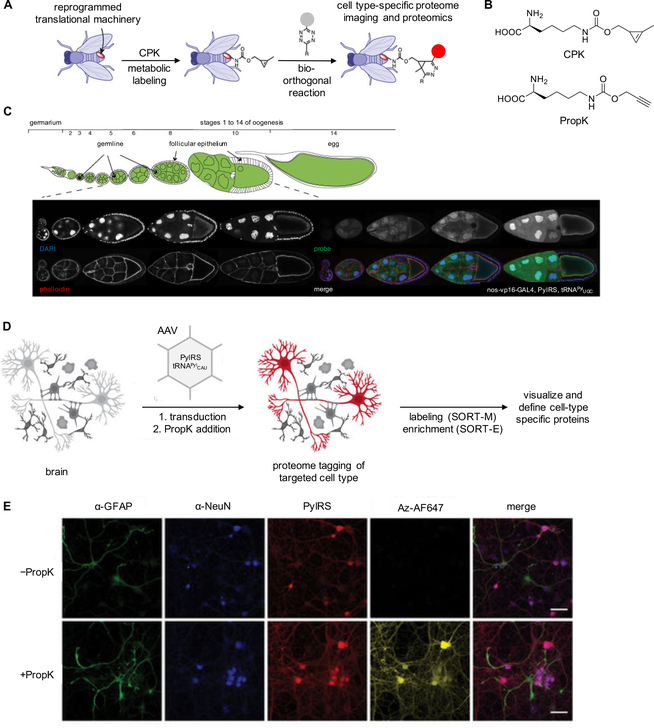
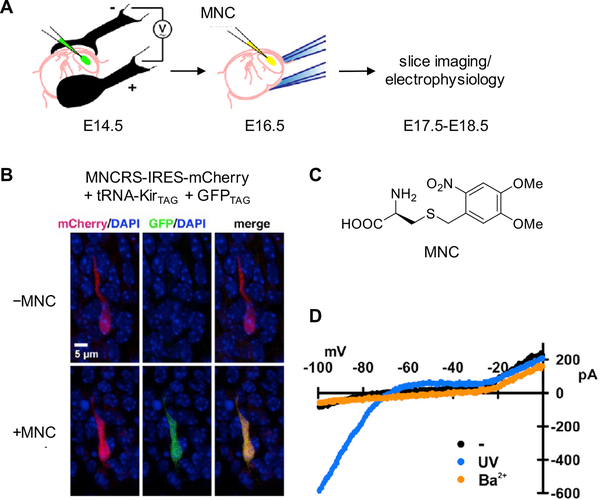
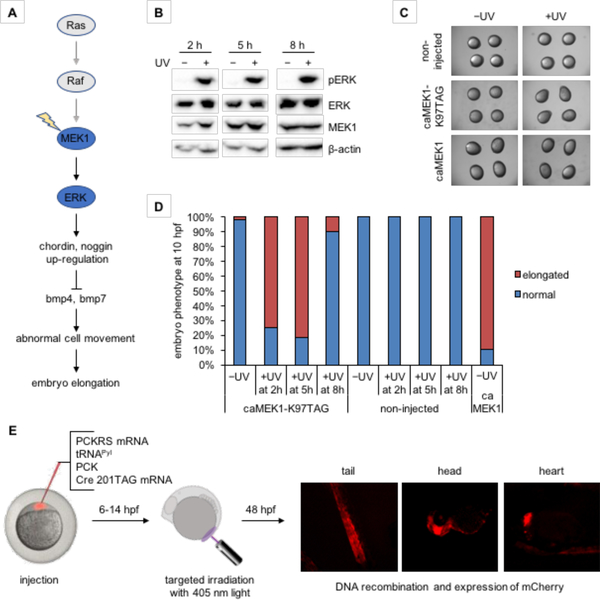
References
-
- Heckler TG; Zama Y; Naka T; Hecht SM, Dipeptide formation with misacylated tRNAPhes. J. Biol. Chem 1983, 258, 4492–4495. - PubMed
-
- Noren CJ; Anthonycahill SJ; Griffith MC; Schultz PG, A General-method for site-specific incorporation of unnatural amino-acids into proteins. Science 1989, 244, 182–188. - PubMed
-
- Ibba M; Hennecke H, Relaxing the substrate specificity of an aminoacyl-tRNA synthetase allows in vitro and in vivo synthesis of proteins containing unnatural amino acids. FEBS Lett. 1995, 364, 272–275. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
