

The Diverse Roles of microRNAs at the Host⁻Virus Interface
- PMID: 30126238
- PMCID: PMC6116274
- DOI: 10.3390/v10080440
The Diverse Roles of microRNAs at the Host⁻Virus Interface
Abstract
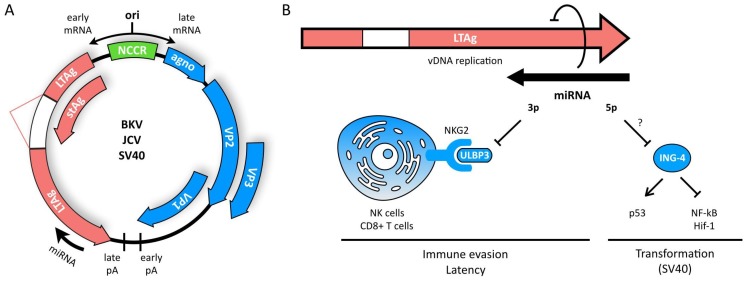
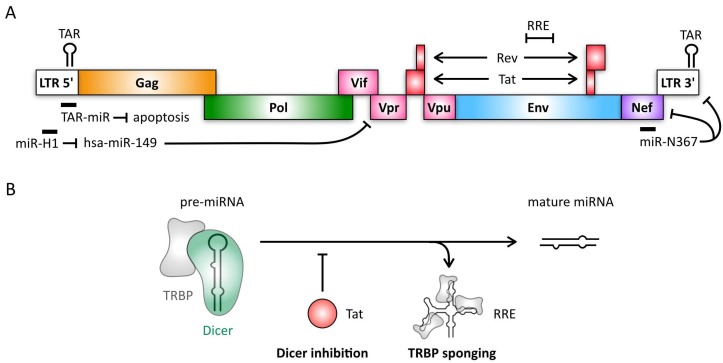
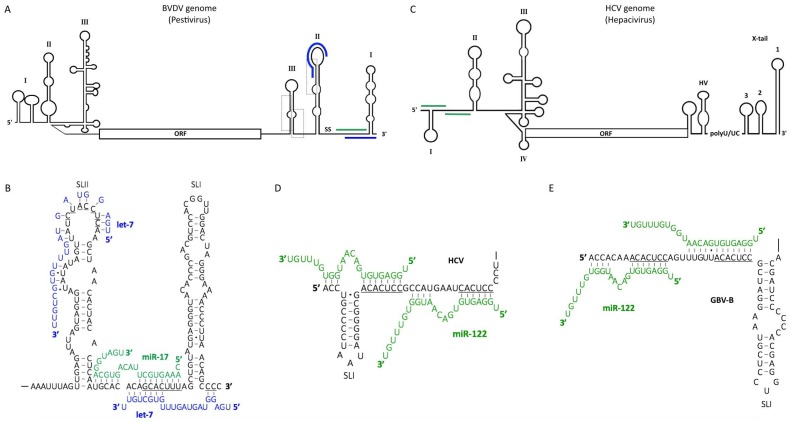
MicroRNAs (miRNAs) are small, non-coding RNAs that regulate gene expression at the post-transcriptional level. Through this activity, they are implicated in almost every cellular process investigated to date. Hence, it is not surprising that miRNAs play diverse roles in regulation of viral infections and antiviral responses. Diverse families of DNA and RNA viruses have been shown to take advantage of cellular miRNAs or produce virally encoded miRNAs that alter host or viral gene expression. MiRNA-mediated changes in gene expression have been demonstrated to modulate viral replication, antiviral immune responses, viral latency, and pathogenesis. Interestingly, viruses mediate both canonical and non-canonical interactions with miRNAs to downregulate specific targets or to promote viral genome stability, translation, and/or RNA accumulation. In this review, we focus on recent findings elucidating several key mechanisms employed by diverse virus families, with a focus on miRNAs at the host⁻virus interface during herpesvirus, polyomavirus, retroviruses, pestivirus, and hepacivirus infections.
Keywords: Herpesviruses; hepaciviruses; immune evasion; latency; microRNAs; pestiviruses; polyomaviruses; retroviruses; viral RNA accumulation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
