

Self-associated molecular patterns mediate cancer immune evasion by engaging Siglecs on T cells
- PMID: 30130255
- PMCID: PMC6205408
- DOI: 10.1172/JCI120612
Self-associated molecular patterns mediate cancer immune evasion by engaging Siglecs on T cells
Abstract
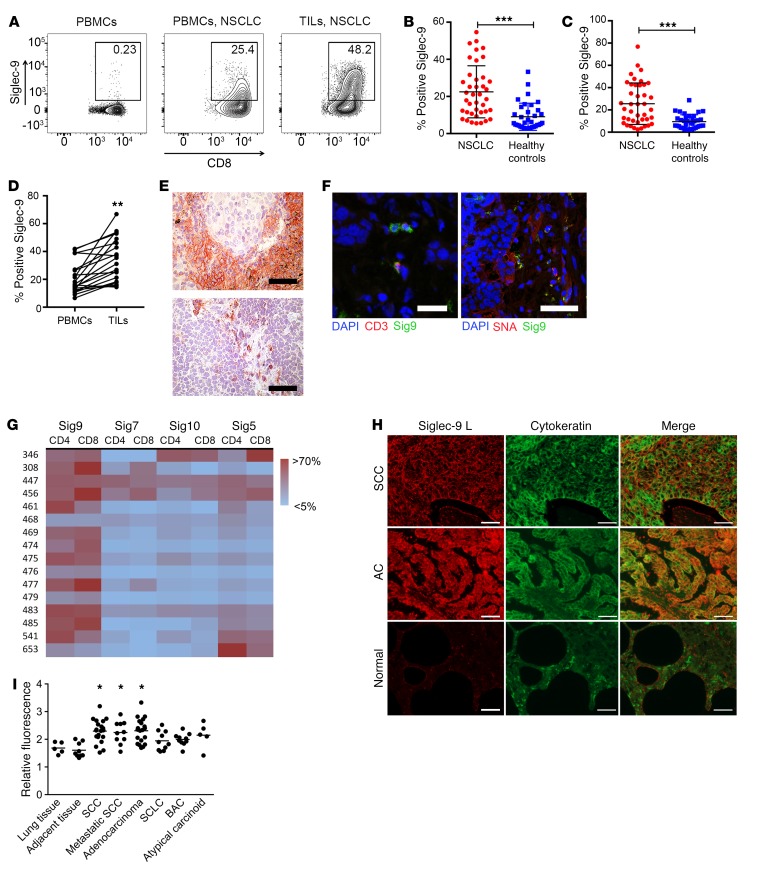
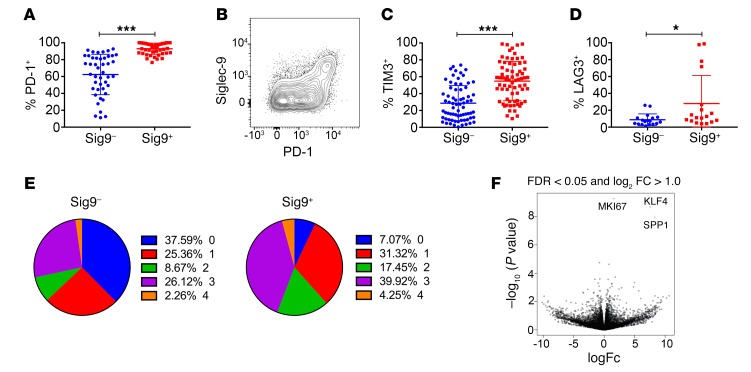
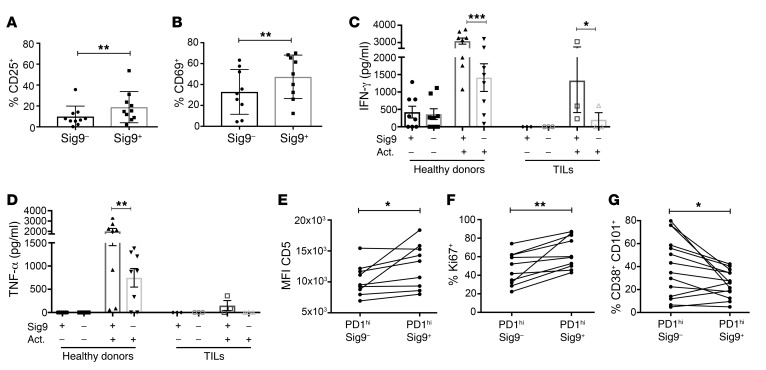
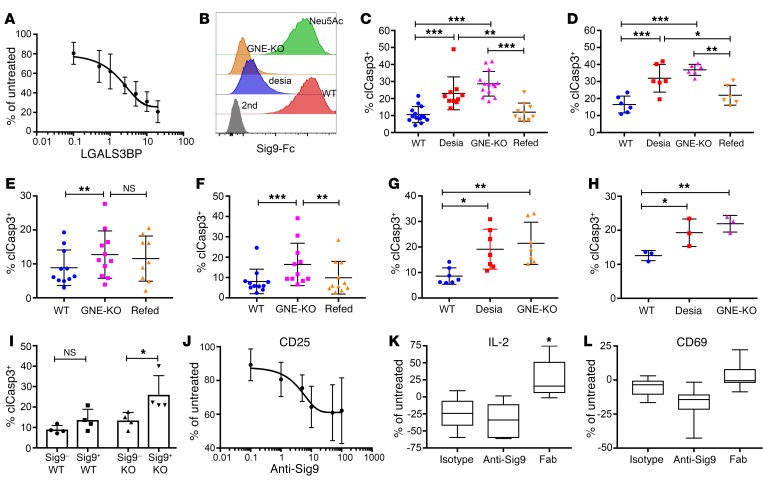
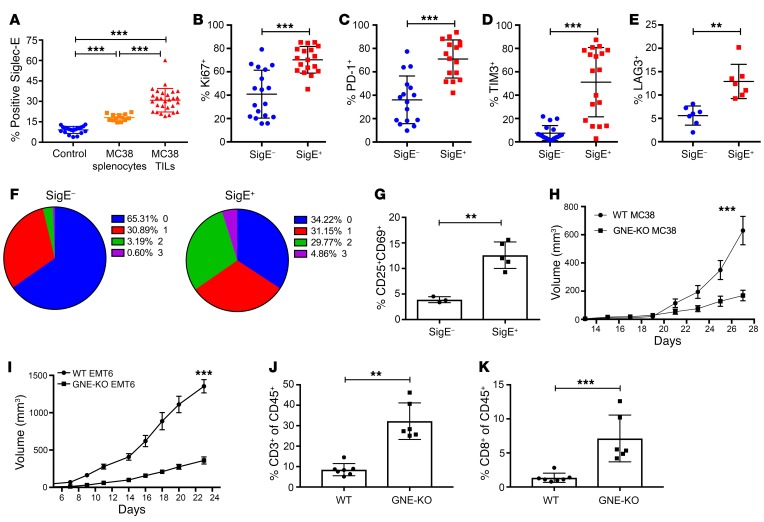
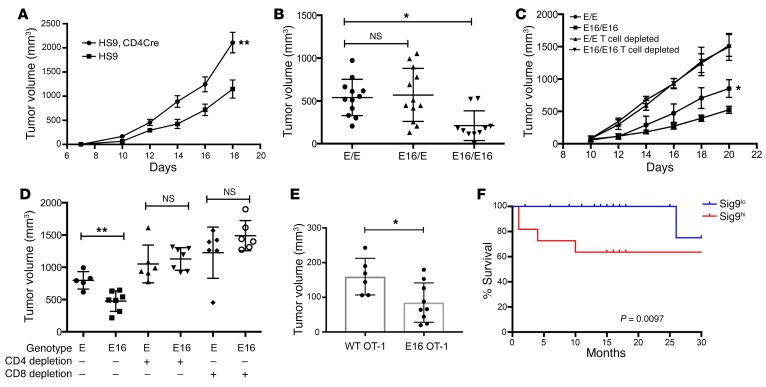
First-generation immune checkpoint inhibitors, including anti-CTLA-4 and anti-programmed death 1 (anti-PD-1) antibodies, have led to major clinical progress, yet resistance frequently leads to treatment failure. Thus, new targets acting on T cells are needed. CD33-related sialic acid-binding immunoglobulin-like lectins (Siglecs) are pattern-recognition immune receptors binding to a range of sialoglycan ligands, which appear to function as self-associated molecular patterns (SAMPs) that suppress autoimmune responses. Siglecs are expressed at very low levels on normal T cells, and these receptors were not until recently considered as interesting targets on T cells for cancer immunotherapy. Here, we show an upregulation of Siglecs, including Siglec-9, on tumor-infiltrating T cells from non-small cell lung cancer (NSCLC), colorectal, and ovarian cancer patients. Siglec-9-expressing T cells coexpressed several inhibitory receptors, including PD-1. Targeting of the sialoglycan-SAMP/Siglec pathway in vitro and in vivo resulted in increased anticancer immunity. T cell expression of Siglec-9 in NSCLC patients correlated with reduced survival, and Siglec-9 polymorphisms showed association with the risk of developing lung and colorectal cancer. Our data identify the sialoglycan-SAMP/Siglec pathway as a potential target for improving T cell activation for immunotherapy.
Keywords: Cancer immunotherapy; Immunology; Oncology; T cells.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
