

Plasma membrane cholesterol trafficking in steroidogenesis
- PMID: 30133326
- PMCID: PMC6988844
- DOI: 10.1096/fj.201800697RRR
Plasma membrane cholesterol trafficking in steroidogenesis
Abstract
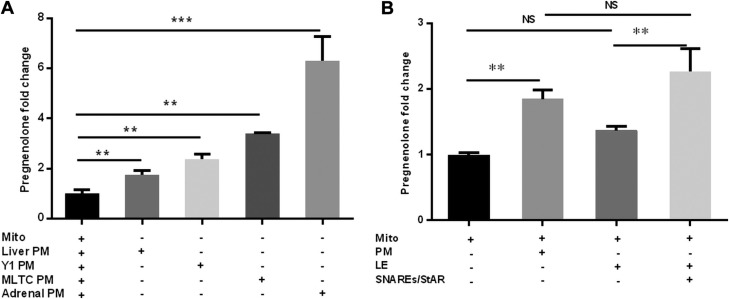
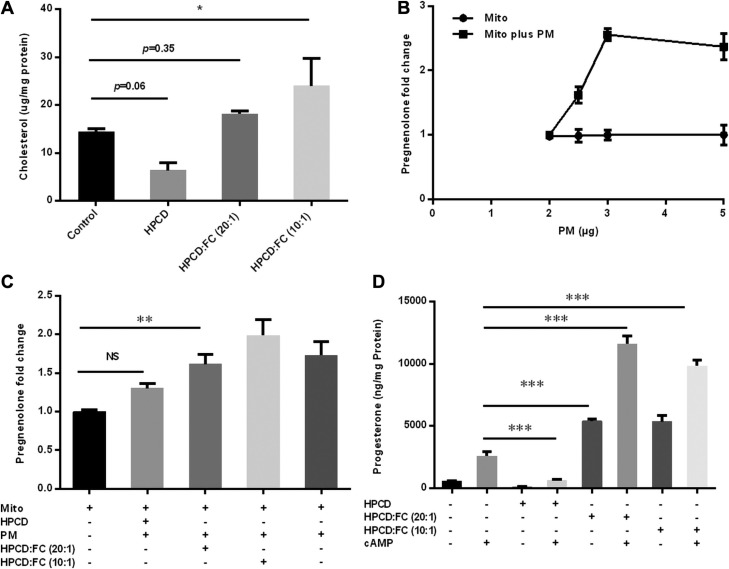
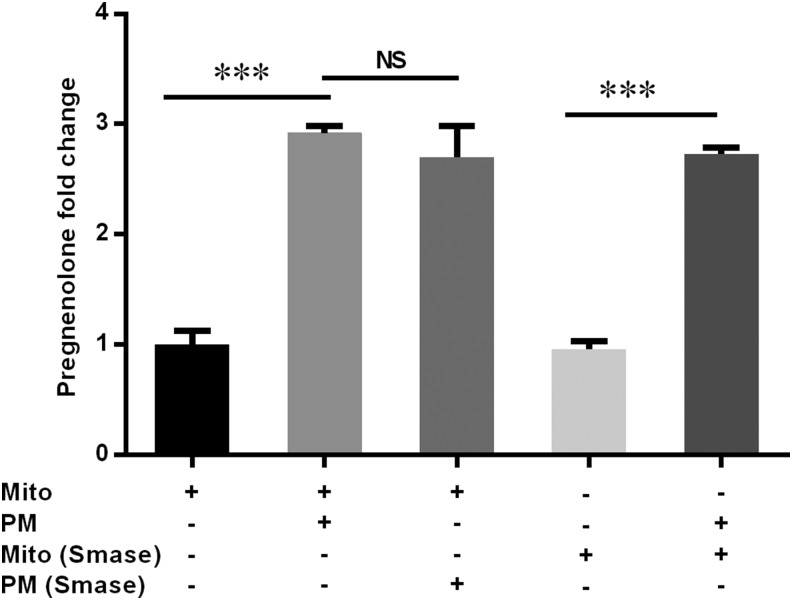
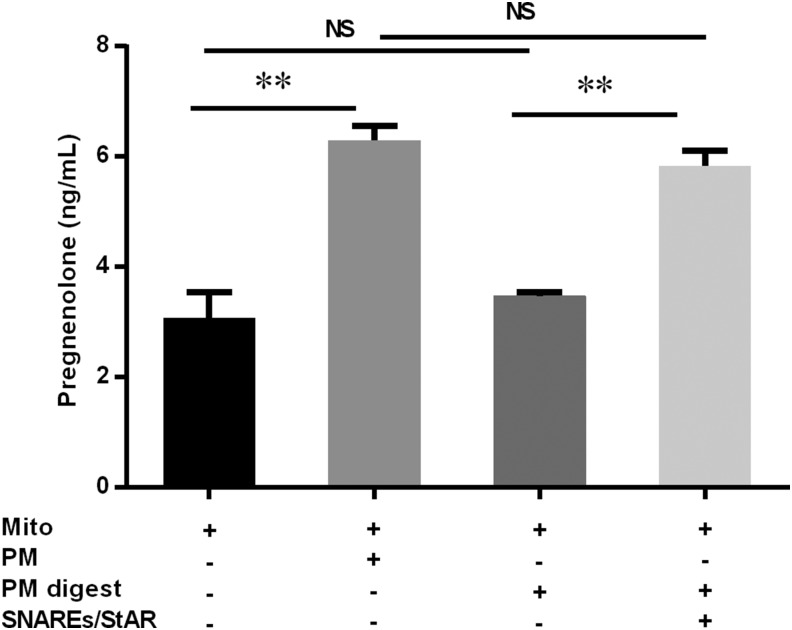
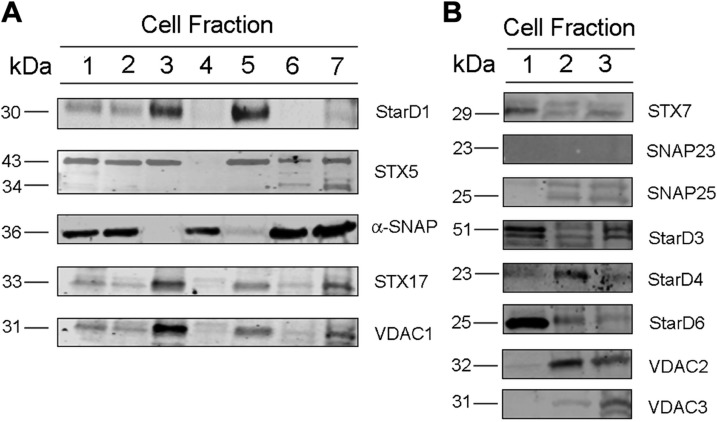
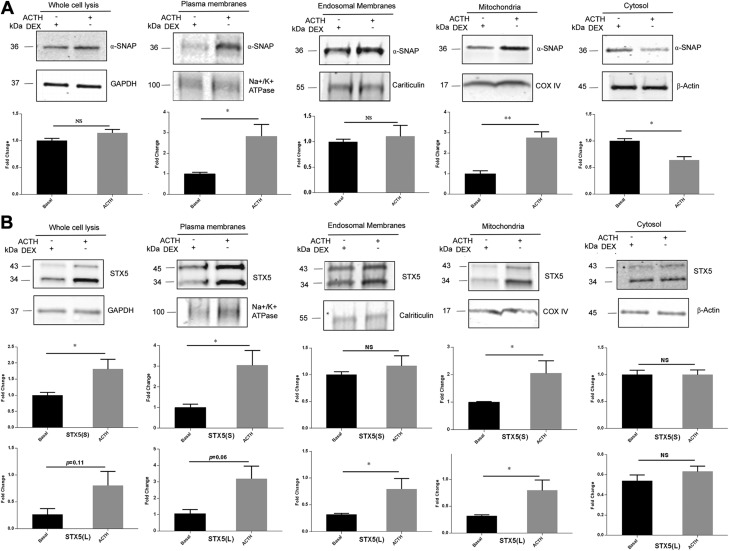
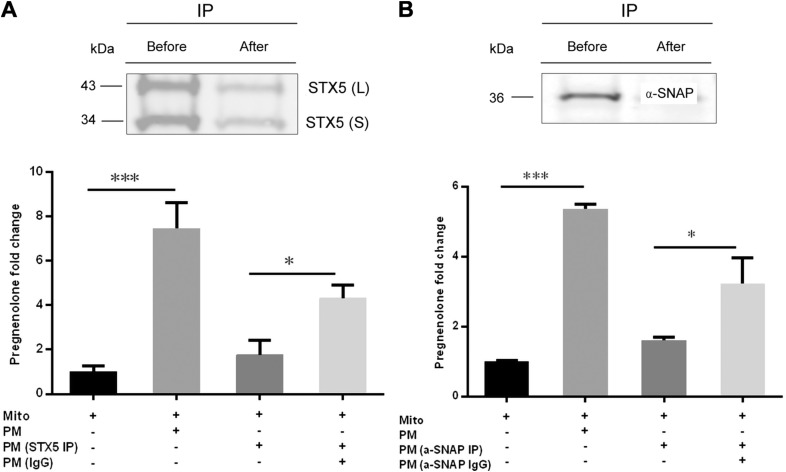
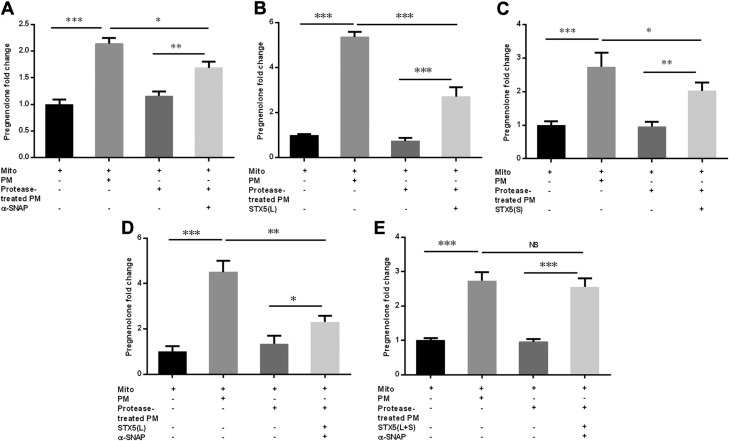
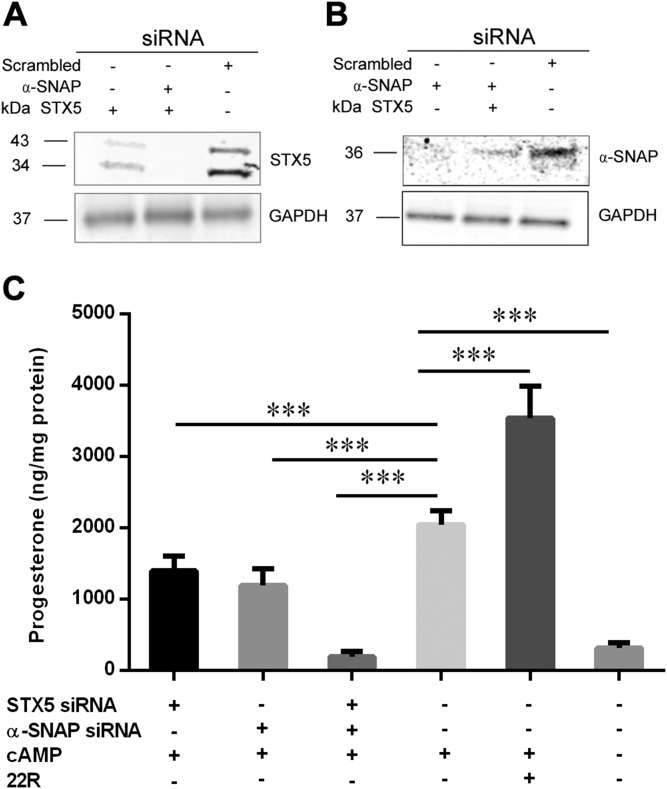
Cholesterol is an important component of plasma membranes (PMs) and the precursor of all steroid hormones. In steroidogenic tissues, upon hormone stimulation, there is a rapid transfer of cholesterol to the mitochondria, which is the site of the initial step in steroidogenesis. In the current study, we examined PM cholesterol trafficking for steroidogenesis. In a mitochondrial reconstitution assay, adrenal PMs supported steroidogenesis in the absence of additional transport proteins. Depletion of cholesterol in PMs by 50% eliminated the membranes' ability to support steroidogenesis in vitro and reduced steroid production in intact Y1 adrenocortical cells. Syntaxin (STX)-5 and α-soluble N-ethylmaleimide-sensitive factor attachment protein (α-SNAP) are enriched in adrenal PMs, and adrenocorticotropic hormone treatment of rats recruited STX5 and α-SNAP to adrenal PMs and mitochondria. Immunodepletion of STX5 and α-SNAP from PMs decreased steroidogenesis supported by PMs in vitro. Protease digestion of PMs decreased, whereas recombinant STX5 or α-SNAP restored, the PMs' ability to support steroidogenesis. Knockdown of either STX5 or α-SNAP in Y1 cells decreased stimulated steroidogenesis. These results indicate that STX5 and α-SNAP facilitate cholesterol trafficking from PMs to mitochondria for adrenal steroid synthesis and underscore the importance of vesicular trafficking of PM cholesterol for steroidogenesis.-Deng, B., Shen, W.-J., Dong, D., Azhar, S., Kraemer, F. B. Plasma membrane cholesterol trafficking in steroidogenesis.
Keywords: SNARE proteins; adrenal gland; mitochondria.
Conflict of interest statement
This work was supported by Merit Review Awards I01BX001923 (to S.A.) and I01BX000398 (to F.B.K.); and Senior Research Career Scientist Award IK6B004200 (to S.A.) from the U.S. Department of Veterans Affairs, Biomedical Laboratory Research Development Program. The authors declare no conflicts of interest.
Figures
References
-
- Lindzey J., Korach K. S. (1997) Steroid hormones. In Endocrinology: Basic and Clinical Principles (Conn P. M., and Melmed S., eds.), pp. 47–62, Humana Press, Totowa, NJ, USA: 10.1007/978-1-59259-641-6_4 - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
