

Peroxisome Proliferator-Activated Receptor γ2 Controls the Rate of Adipose Tissue Lipid Storage and Determines Metabolic Flexibility
- PMID: 30134163
- PMCID: PMC6113930
- DOI: 10.1016/j.celrep.2018.07.063
Peroxisome Proliferator-Activated Receptor γ2 Controls the Rate of Adipose Tissue Lipid Storage and Determines Metabolic Flexibility
Abstract
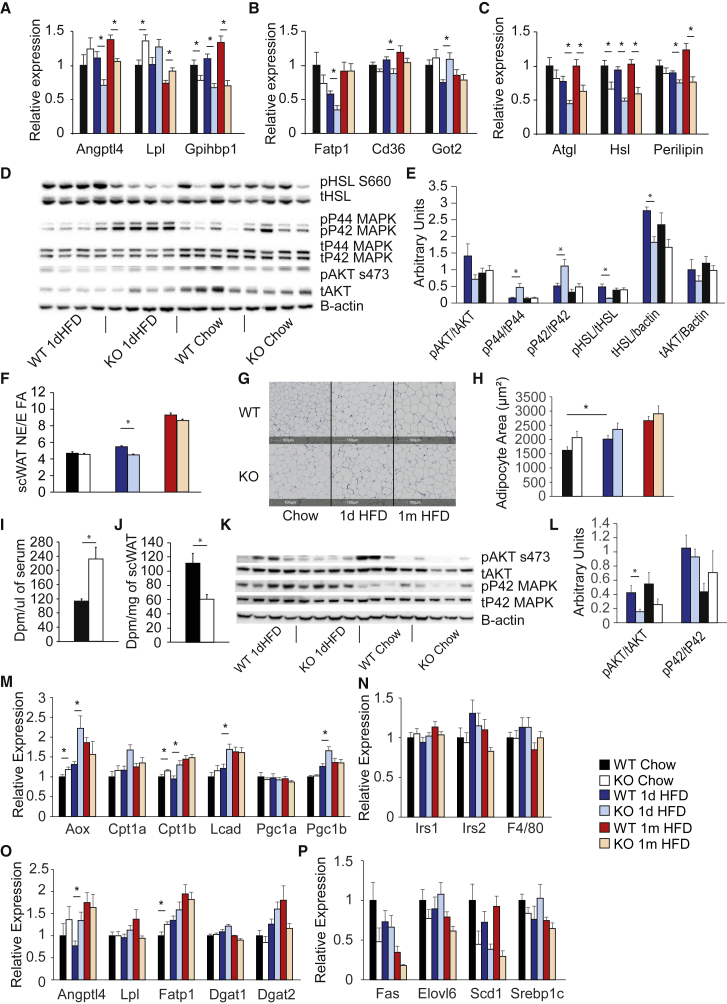
One understudied function of white adipose tissue (AT) is its role in postprandial lipid buffering. In this study, we demonstrate that mice lacking the adipose tissue-specific transcription factor peroxisome proliferator-activated receptor γ2 (PPARγ2) exhibit a defect in their rate of adipose tissue lipid storage. Impaired adipose tissue storage rate reduces metabolic flexibility, without compromising fasted glucose tolerance or insulin sensitivity, even following prolonged high-fat feeding. However, acutely overfeeding PPARγ2-KO mice caused a 10-fold increase in insulin levels compared with controls. Although impaired adipose tissue storage rate did not result in insulin resistance in young mice, 1-year-old PPARγ2-KO mice developed skeletal muscle insulin resistance. Our data indicate that failed adipose tissue storage may occur prior to defects in glucose handling and that overfeeding protocols may uncover genes controlling adipose tissue storage rate, as opposed to capacity, and act as a diagnostic test for early-stage human metabolic disease.
Keywords: PPAR; PPARγ; PPARγ2; Randle; WAT; adipose tissue; insulin resistance; metabolic flexibility; overfeeding; overnutrition.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Figures




References
-
- Danforth E., Jr. Failure of adipocyte differentiation causes type II diabetes mellitus? Nat. Genet. 2000;26:13. - PubMed
-
- Escher P., Braissant O., Basu-Modak S., Michalik L., Wahli W., Desvergne B. Rat PPARs: quantitative analysis in adult rat tissues and regulation in fasting and refeeding. Endocrinology. 2001;142:4195–4202. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MC_G0802535/MRC_/Medical Research Council/United Kingdom
- MC_UU_12012/2/MRC_/Medical Research Council/United Kingdom
- MC_UU_00014/5/MRC_/Medical Research Council/United Kingdom
- MC_UU_00014/2/MRC_/Medical Research Council/United Kingdom
- G0600717/MRC_/Medical Research Council/United Kingdom
- WT_/Wellcome Trust/United Kingdom
- G0802051/MRC_/Medical Research Council/United Kingdom
- MC_UU_12012/5/MRC_/Medical Research Council/United Kingdom
- G0400192/MRC_/Medical Research Council/United Kingdom
- RG/12/13/29853/BHF_/British Heart Foundation/United Kingdom
- RG/18/7/33636/BHF_/British Heart Foundation/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
