

Morphological intelligence counters foot slipping in the desert locust and dynamic robots
- PMID: 30135101
- PMCID: PMC6130395
- DOI: 10.1073/pnas.1804239115
Morphological intelligence counters foot slipping in the desert locust and dynamic robots
Abstract
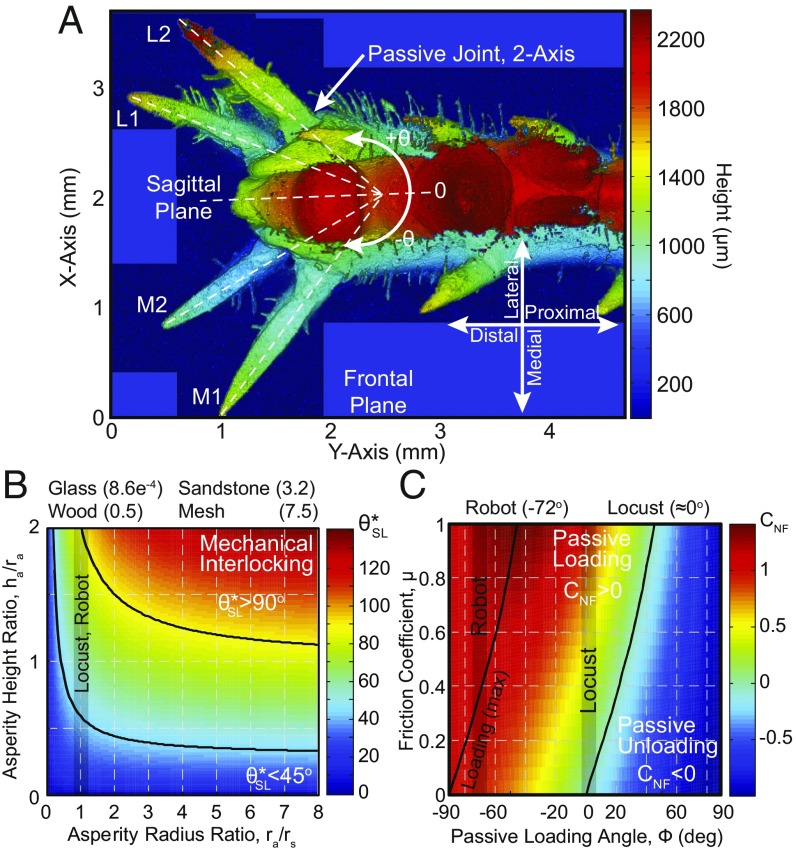
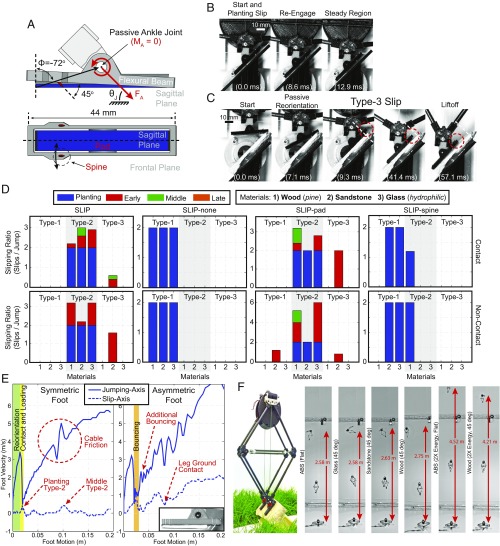
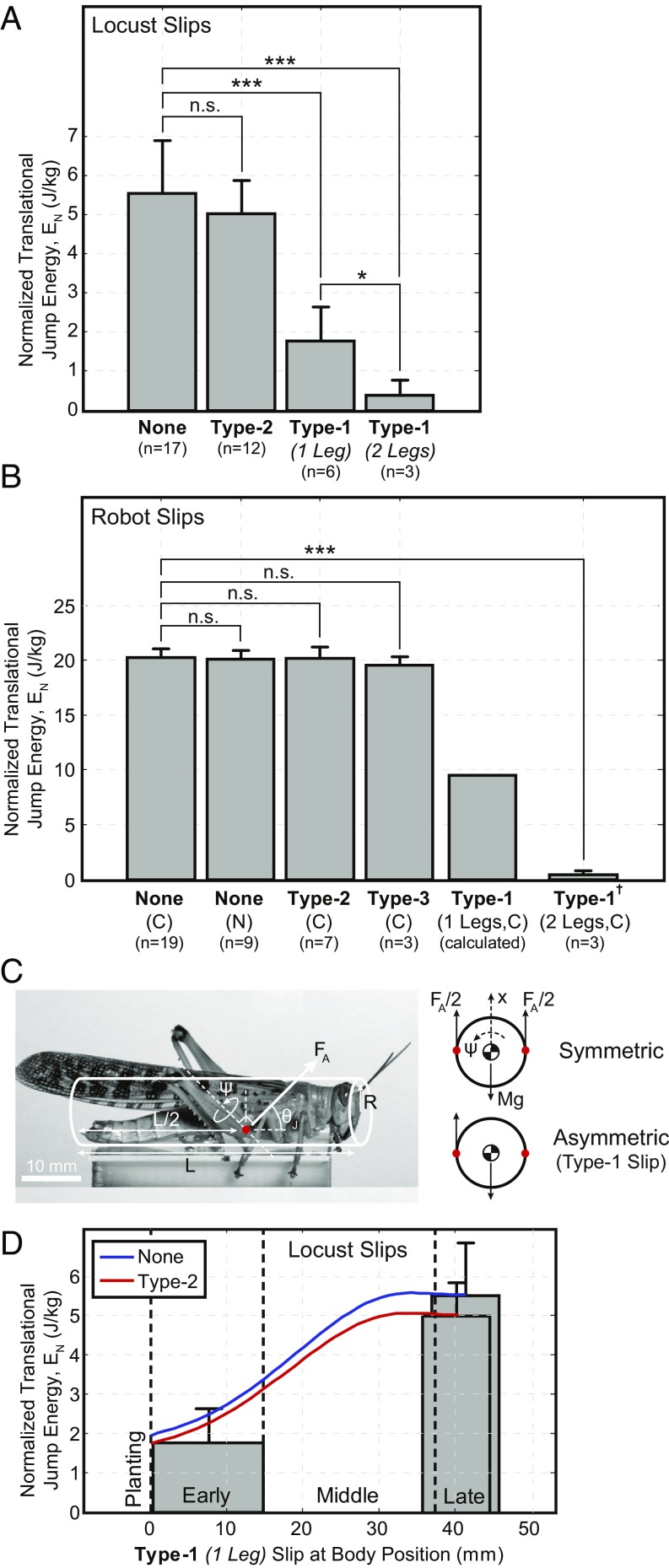
During dynamic terrestrial locomotion, animals use complex multifunctional feet to extract friction from the environment. However, whether roboticists assume sufficient surface friction for locomotion or actively compensate for slipping, they use relatively simple point-contact feet. We seek to understand and extract the morphological adaptations of animal feet that contribute to enhancing friction on diverse surfaces, such as the desert locust (Schistocerca gregaria) [Bennet-Clark HC (1975) J Exp Biol 63:53-83], which has both wet adhesive pads and spines. A buckling region in their knee to accommodate slipping [Bayley TG, Sutton GP, Burrows M (2012) J Exp Biol 215:1151-1161], slow nerve conduction velocity (0.5-3 m/s) [Pearson KG, Stein RB, Malhotra SK (1970) J Exp Biol 53:299-316], and an ecological pressure to enhance jumping performance for survival [Hawlena D, Kress H, Dufresne ER, Schmitz OJ (2011) Funct Ecol 25:279-288] further suggest that the locust operates near the limits of its surface friction, but without sufficient time to actively control its feet. Therefore, all surface adaptation must be through passive mechanics (morphological intelligence), which are unknown. Here, we report the slipping behavior, dynamic attachment, passive mechanics, and interplay between the spines and adhesive pads, studied through both biological and robotic experiments, which contribute to the locust's ability to jump robustly from diverse surfaces. We found slipping to be surface-dependent and common (e.g., wood 1.32 ± 1.19 slips per jump), yet the morphological intelligence of the feet produces a significant chance to reengage the surface (e.g., wood 1.10 ± 1.13 reengagements per jump). Additionally, a discovered noncontact-type jump, further studied robotically, broadens the applicability of the morphological adaptations to both static and dynamic attachment.
Keywords: friction; jump; locust; robot; slip.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Dickinson MH, et al. How animals move: An integrative view. Science. 2000;288:100–106. - PubMed
-
- Noh M, Kim SW, An S, Koh JS, Cho KJ. Flea-inspired catapult mechanism for miniature jumping robots. IEEE Trans Rob. 2012;28:1007–1018.
-
- Kovac M, Fuchs M, Guignard A, Zufferey JC, Floreano D. A miniature 7g jumping robot. IEEE International Conference on Robotics and Automation. 2008:373–378.
-
- Beck A, et al. Jump stabilization and landing control by wing-spreading of a locust-inspired jumper. Bioinspiration Biomimetics. 2017;12:066006. - PubMed
-
- Zhao J, et al. MSU jumper: A single-motor-actuated miniature steerable jumping robot. IEEE Trans Rob. 2013;29:602–614.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
