

Heterogeneity of hypochlorous acid production in individual neutrophil phagosomes revealed by a rhodamine-based probe
- PMID: 30135208
- PMCID: PMC6177602
- DOI: 10.1074/jbc.RA118.004789
Heterogeneity of hypochlorous acid production in individual neutrophil phagosomes revealed by a rhodamine-based probe
Abstract
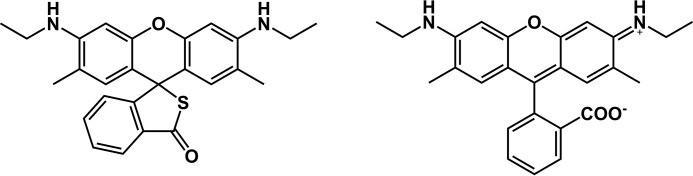
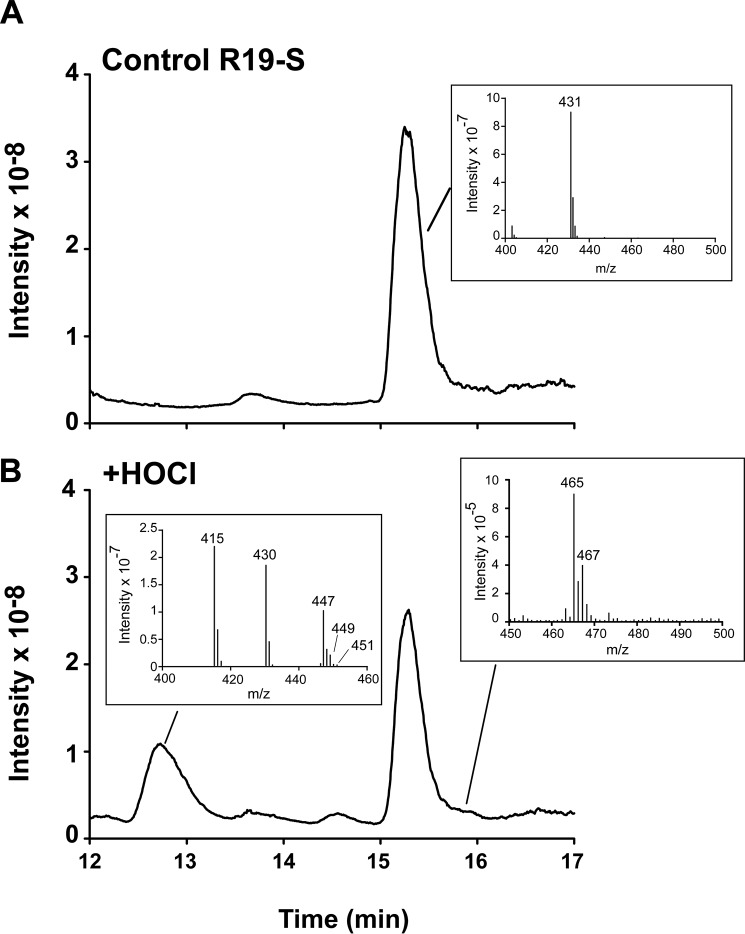
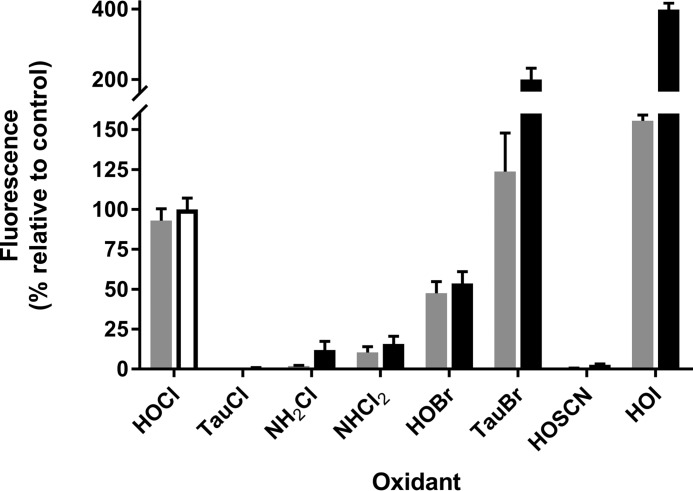
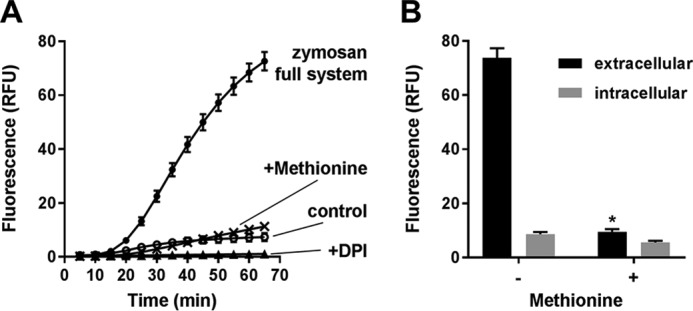
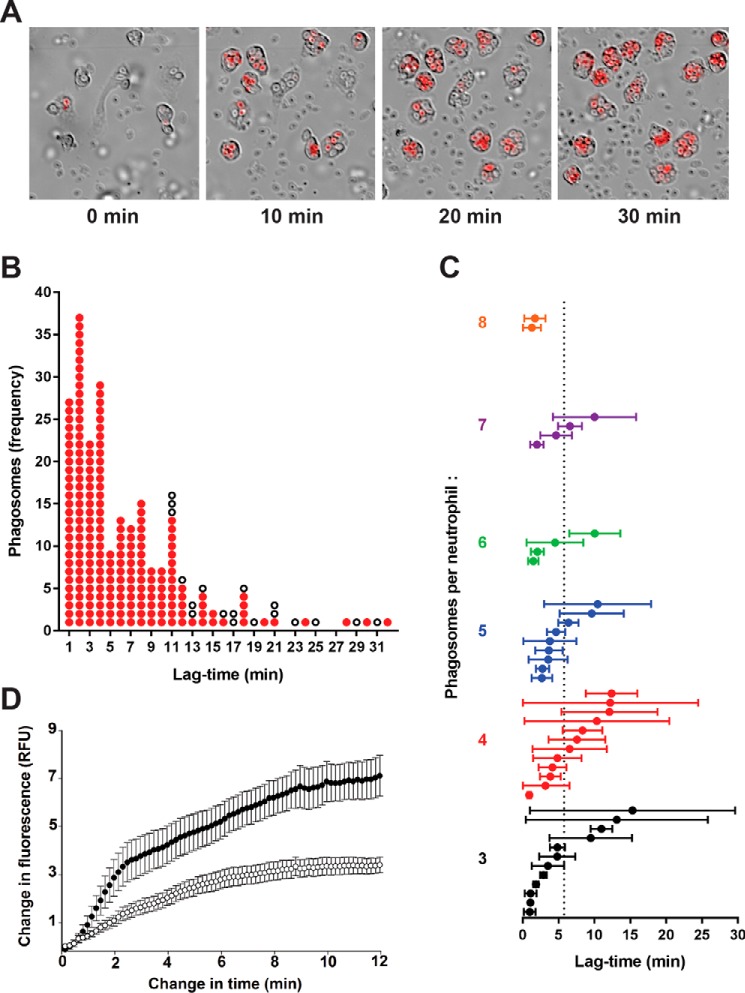
The rhodamine-based probe R19-S has been shown to react with hypochlorous acid (HOCl) to yield fluorescent R19, but not with some other oxidants including hydrogen peroxide. Here, we further examined the specificity of R19-S and used it for real-time monitoring of HOCl production in neutrophil phagosomes. We show that it also reacts rapidly with hypobromous acid, bromamines, and hypoiodous acid, indicating that R19-S responds to these reactive halogen species as well as HOCl. Hypothiocyanous acid and taurine chloramine were unreactive, however, and ammonia chloramine and dichloramine reacted only very slowly. MS analyses revealed additional products from the reaction of HOCl with R19-S, including a chlorinated species as a minor product. Of note, phagocytosis of opsonized zymosan or Staphylococcus aureus by neutrophils was accompanied by an increase in R19 fluorescence. This increase depended on NADPH oxidase and myeloperoxidase activities, and detection of chlorinated R19-S confirmed its specificity for HOCl. Using live-cell imaging to track individual phagosomes in single neutrophils, we observed considerable heterogeneity among the phagosomes in the time from ingestion of a zymosan particle to when fluorescence was first detected, ranging from 1 to >30 min. However, once initiated, the subsequent fluorescence increase was uniform, reaching a similar maximum in ∼10 min. Our results confirm the utility of R19-S for detecting HOCl in real-time and provide definitive evidence that isolated neutrophils produce HOCl in phagosomes. The intriguing variability in the onset of HOCl production among phagosomes identified here could influence the way they kill ingested bacteria.
Keywords: host-pathogen interaction; hypobromous; hypochlorous; myeloperoxidase; neutrophil; oxidative stress; phagocytosis.
© 2018 Albrett et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures



References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
