

Enterovirus D68 infection induces IL-17-dependent neutrophilic airway inflammation and hyperresponsiveness
- PMID: 30135310
- PMCID: PMC6141171
- DOI: 10.1172/jci.insight.121882
Enterovirus D68 infection induces IL-17-dependent neutrophilic airway inflammation and hyperresponsiveness
Abstract
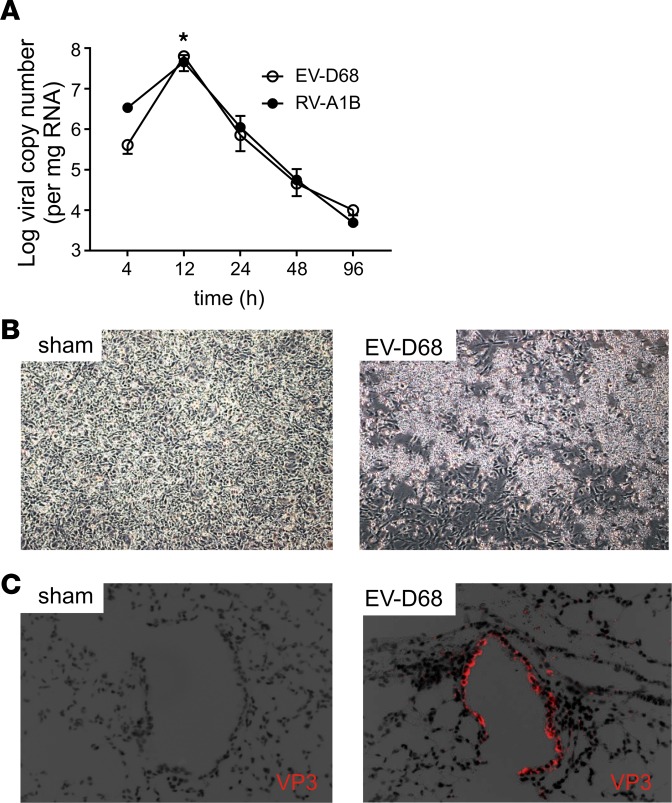
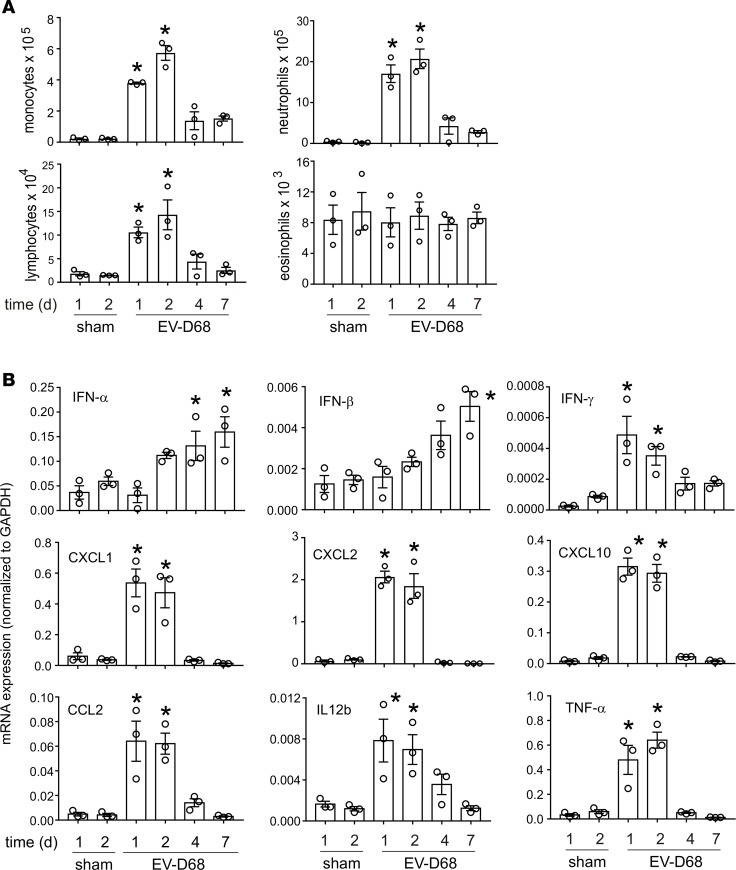
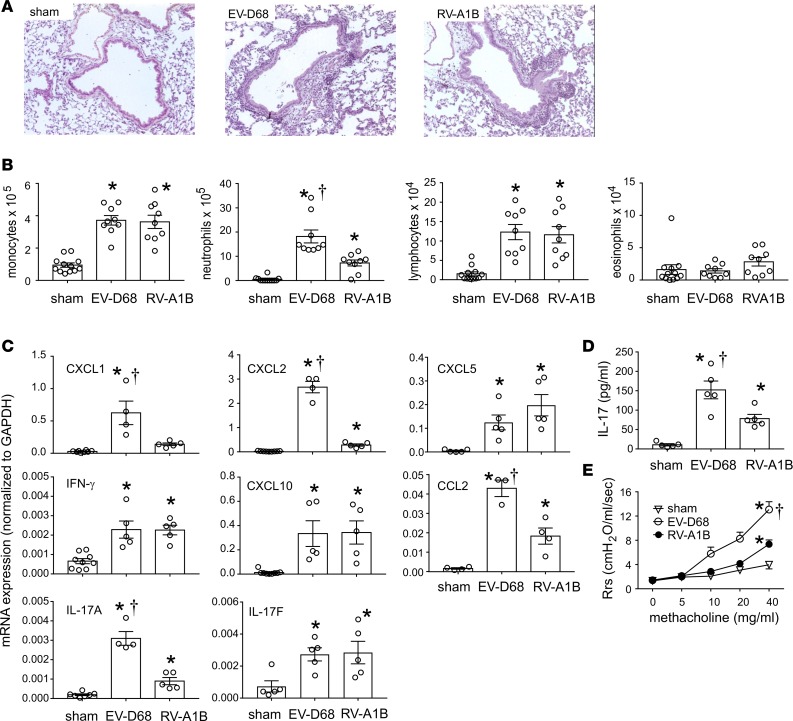
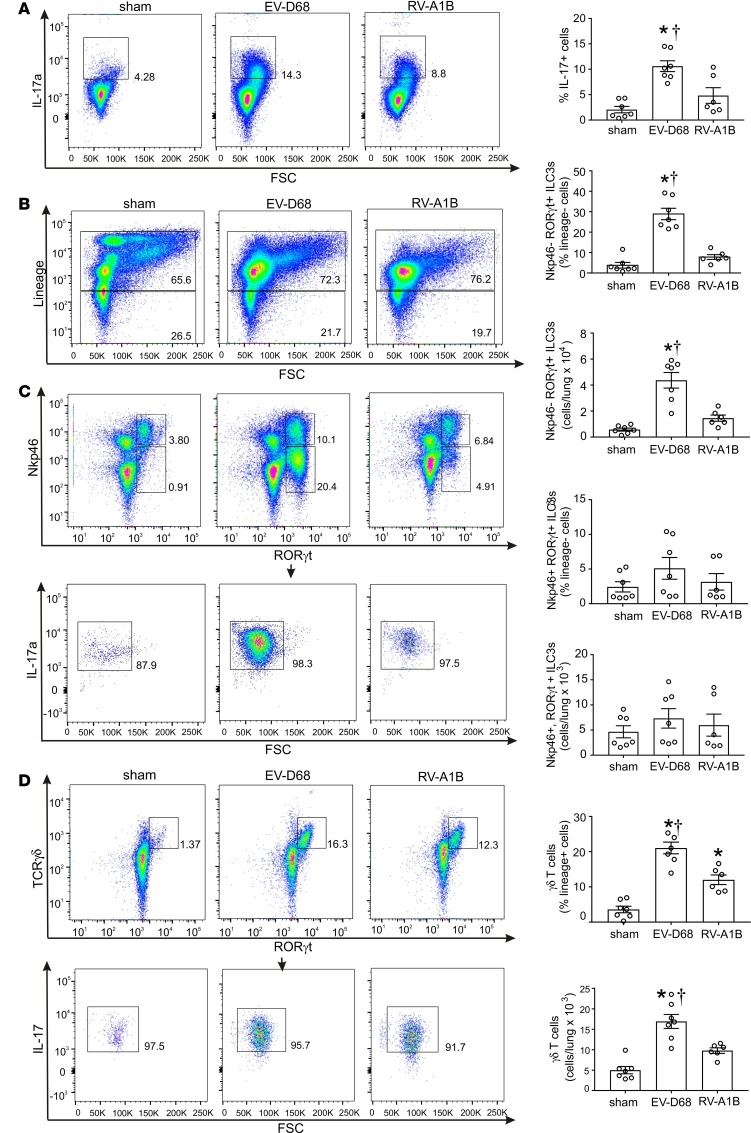
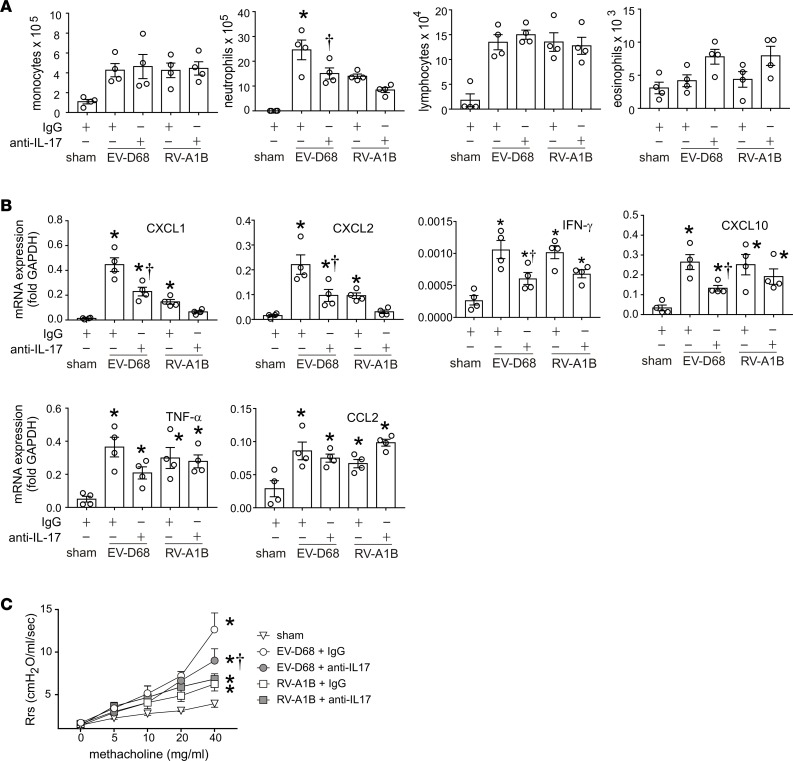
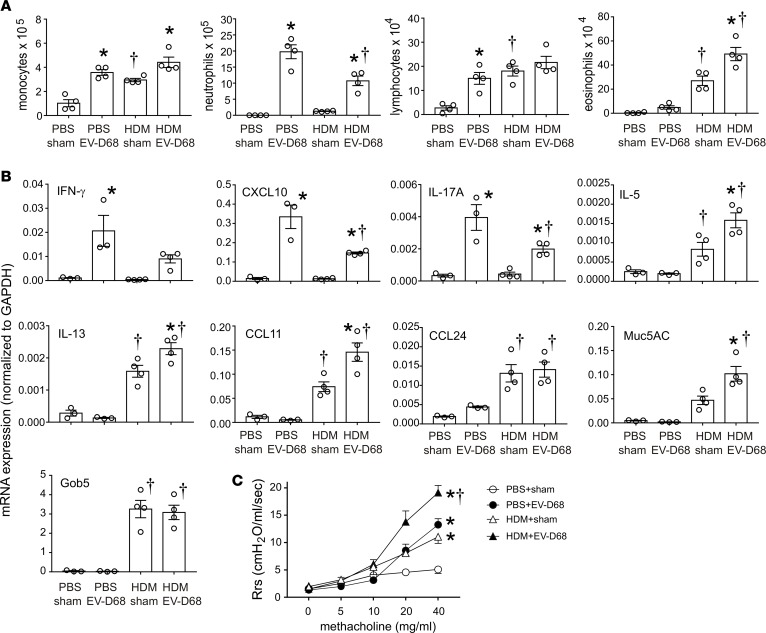
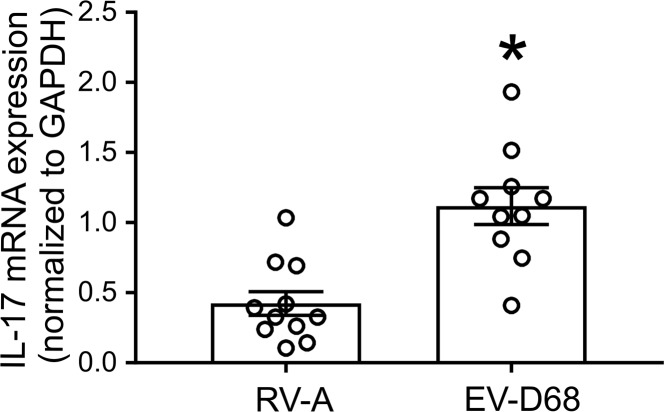
Enterovirus D68 (EV-D68) shares biologic features with rhinovirus (RV). In 2014, a nationwide outbreak of EV-D68 was associated with severe asthma-like symptoms. We sought to develop a mouse model of EV-D68 infection and determine the mechanisms underlying airway disease. BALB/c mice were inoculated intranasally with EV-D68 (2014 isolate), RV-A1B, or sham, alone or in combination with anti-IL-17A or house dust mite (HDM) treatment. Like RV-A1B, lung EV-D68 viral RNA peaked 12 hours after infection. EV-D68 induced airway inflammation, expression of cytokines (TNF-α, IL-6, IL-12b, IL-17A, CXCL1, CXCL2, CXCL10, and CCL2), and airway hyperresponsiveness, which were suppressed by anti-IL-17A antibody. Neutrophilic inflammation and airway responsiveness were significantly higher after EV-D68 compared with RV-A1B infection. Flow cytometry showed increased lineage-, NKp46-, RORγt+ IL-17+ILC3s and γδ T cells in the lungs of EV-D68-treated mice compared with those in RV-treated mice. EV-D68 infection of HDM-exposed mice induced additive or synergistic increases in BAL neutrophils and eosinophils and expression of IL-17, CCL11, IL-5, and Muc5AC. Finally, patients from the 2014 epidemic period with EV-D68 showed significantly higher nasopharyngeal IL-17 mRNA levels compared with patients with RV-A infection. EV-D68 infection induces IL-17-dependent airway inflammation and hyperresponsiveness, which is greater than that generated by RV-A1B, consistent with the clinical picture of severe asthma-like symptoms.
Keywords: Allergy; Asthma; Cytokines; Infectious disease; Pulmonology.
Conflict of interest statement
Figures
References
-
- Oberste MS, et al. Enterovirus 68 is associated with respiratory illness and shares biological features with both the enteroviruses and the rhinoviruses. J Gen Virol. 2004;85(Pt 9):2577–2584. - PubMed
-
- Centers for Disease Control Prevention (CDC) Clusters of acute respiratory illness associated with human enterovirus 68--Asia, Europe, and United States, 2008-2010. MMWR Morb Mortal Wkly Rep. 2011;60(38):1301–1304. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
