

Calcium imaging and dynamic causal modelling reveal brain-wide changes in effective connectivity and synaptic dynamics during epileptic seizures
- PMID: 30138336
- PMCID: PMC6124808
- DOI: 10.1371/journal.pcbi.1006375
Calcium imaging and dynamic causal modelling reveal brain-wide changes in effective connectivity and synaptic dynamics during epileptic seizures
Abstract
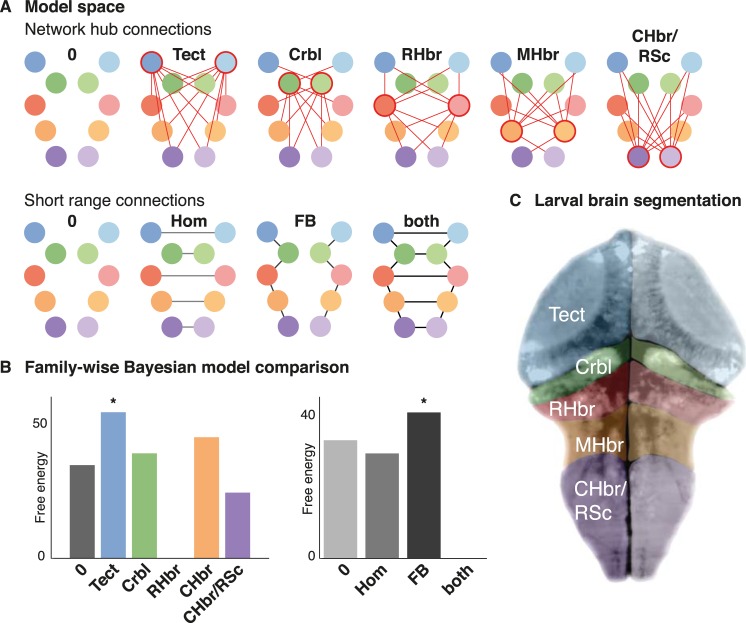
Pathophysiological explanations of epilepsy typically focus on either the micro/mesoscale (e.g. excitation-inhibition imbalance), or on the macroscale (e.g. network architecture). Linking abnormalities across spatial scales remains difficult, partly because of technical limitations in measuring neuronal signatures concurrently at the scales involved. Here we use light sheet imaging of the larval zebrafish brain during acute epileptic seizure induced with pentylenetetrazole. Spectral changes of spontaneous neuronal activity during the seizure are then modelled using neural mass models, allowing Bayesian inference on changes in effective network connectivity and their underlying synaptic dynamics. This dynamic causal modelling of seizures in the zebrafish brain reveals concurrent changes in synaptic coupling at macro- and mesoscale. Fluctuations of both synaptic connection strength and their temporal dynamics are required to explain observed seizure patterns. These findings highlight distinct changes in local (intrinsic) and long-range (extrinsic) synaptic transmission dynamics as a possible seizure pathomechanism and illustrate how our Bayesian model inversion approach can be used to link existing neural mass models of seizure activity and novel experimental methods.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
-
- Depaulis A, David O, Charpier S. The genetic absence epilepsy rat from Strasbourg as a model to decipher the neuronal and network mechanisms of generalized idiopathic epilepsies. J Neurosci Methods. 2015;:1–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
