

Parcellating Cerebral Cortex: How Invasive Animal Studies Inform Noninvasive Mapmaking in Humans
- PMID: 30138588
- PMCID: PMC6149530
- DOI: 10.1016/j.neuron.2018.07.002
Parcellating Cerebral Cortex: How Invasive Animal Studies Inform Noninvasive Mapmaking in Humans
Abstract
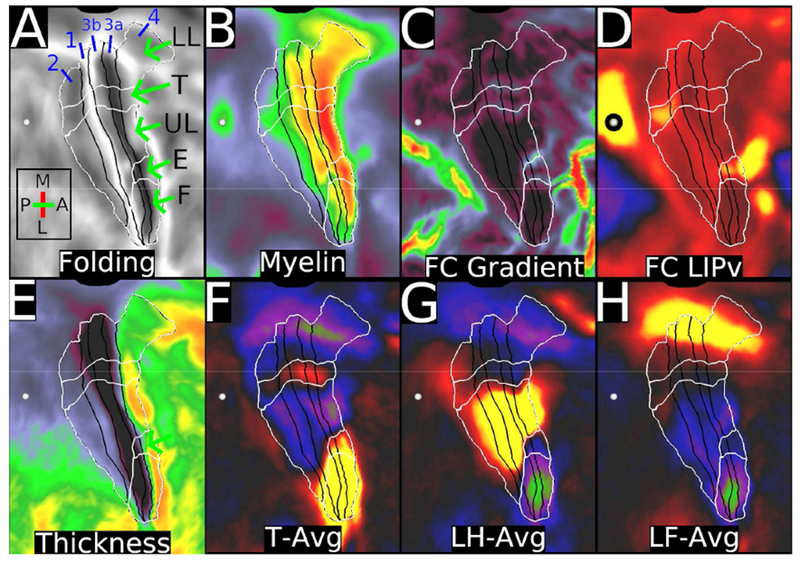
The cerebral cortex in mammals contains a mosaic of cortical areas that differ in function, architecture, connectivity, and/or topographic organization. A combination of local connectivity (within-area microcircuitry) and long-distance (between-area) connectivity enables each area to perform a unique set of computations. Some areas also have characteristic within-area mesoscale organization, reflecting specialized representations of distinct types of information. Cortical areas interact with one another to form functional networks that mediate behavior, and each area may be a part of multiple, partially overlapping networks. Given their importance to the understanding of brain organization, mapping cortical areas across species is a major objective of systems neuroscience and has been a century-long challenge. Here, we review recent progress in multi-modal mapping of mouse and nonhuman primate cortex, mainly using invasive experimental methods. These studies also provide a neuroanatomical foundation for mapping human cerebral cortex using noninvasive neuroimaging, including a new map of human cortical areas that we generated using a semiautomated analysis of high-quality, multimodal neuroimaging data. We contrast our semiautomated approach to human multimodal cortical mapping with various extant fully automated human brain parcellations that are based on only a single imaging modality and offer suggestions on how to best advance the noninvasive brain parcellation field. We discuss the limitations as well as the strengths of current noninvasive methods of mapping brain function, architecture, connectivity, and topography and of current approaches to mapping the brain's functional networks.
Keywords: architectonics; brain; connectivity; cortical areas; function; networks; neuroanatomy; nonhuman primate; parcellation; topography.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures






References
-
- Allman JM, and Kaas JH. (1974) The organization of the second visual area (V II) in the owl monkey: a second order transformation of the visual hemifield. Brain Res 76, 247–265. - PubMed
-
- Allman JM, and Kaas JH. (1975) The dorsomedial cortical visual area: a third tier area in the occipital lobe of the owl monkey (Aotus trivirgatus). Brain Res 100, 473–487. - PubMed
-
- Allman JM, and Kaas JH. (1976) Representation of the visual field on the medial wall of occipital-parietal cortex in the owl monkey. Science. 191, 572–575. - PubMed
-
- Amunts K, Lepage C, Borgeat L, Mohlberg H, Dickscheid T, Rousseau ME, Bludau S, Bazin PL, Lewis LB, Oros-Peusquens AM, Shah NJ, Lippert T, Zilles K, and Evans AC. (2013) BigBrain: an ultrahigh-resolution 3D human brain model. Science. 340, 1472–1475. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
