

A Dynamic Interplay within the Frontoparietal Network Underlies Rhythmic Spatial Attention
- PMID: 30138590
- PMCID: PMC6474777
- DOI: 10.1016/j.neuron.2018.07.038
A Dynamic Interplay within the Frontoparietal Network Underlies Rhythmic Spatial Attention
Abstract
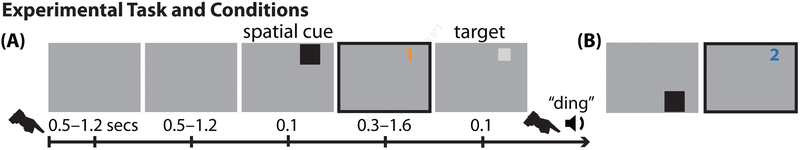
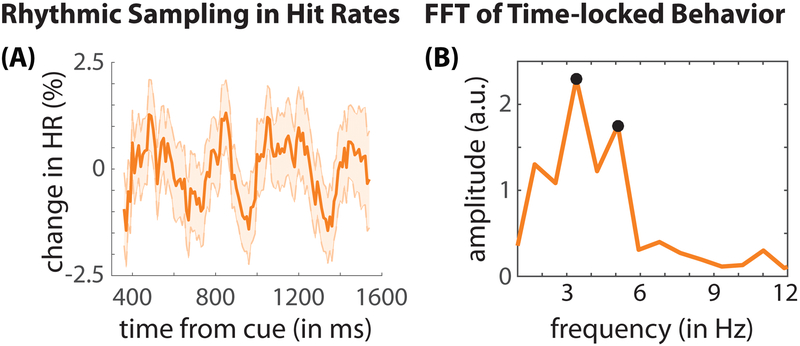
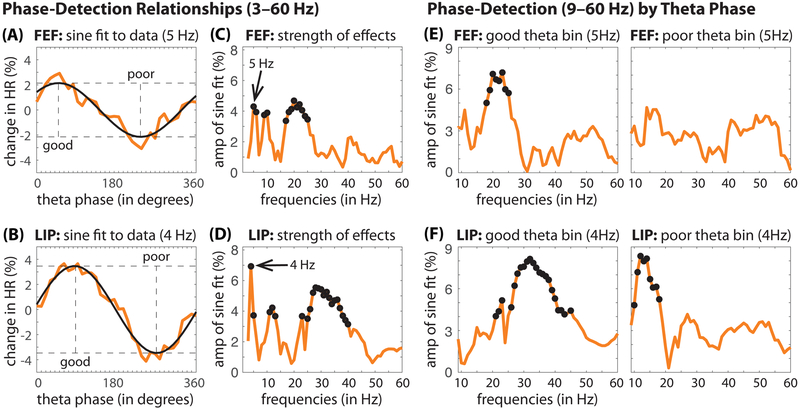
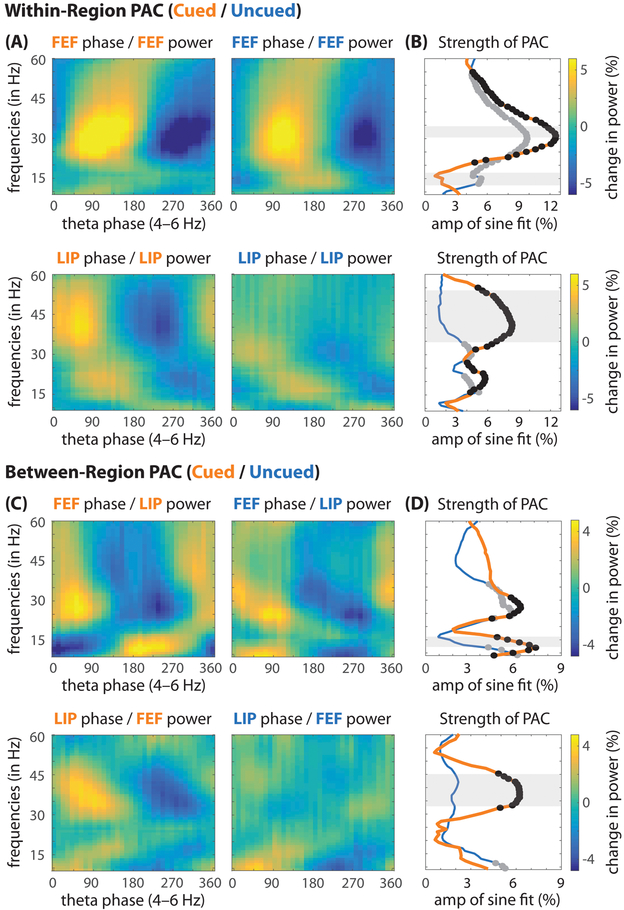
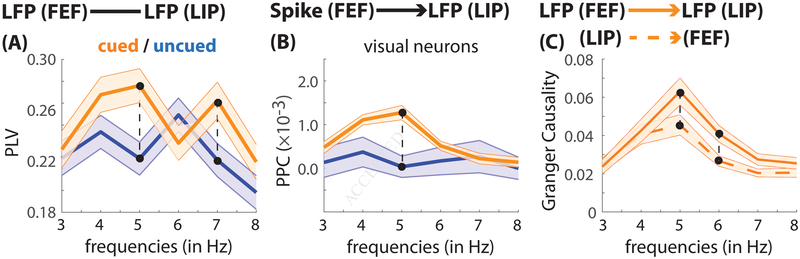
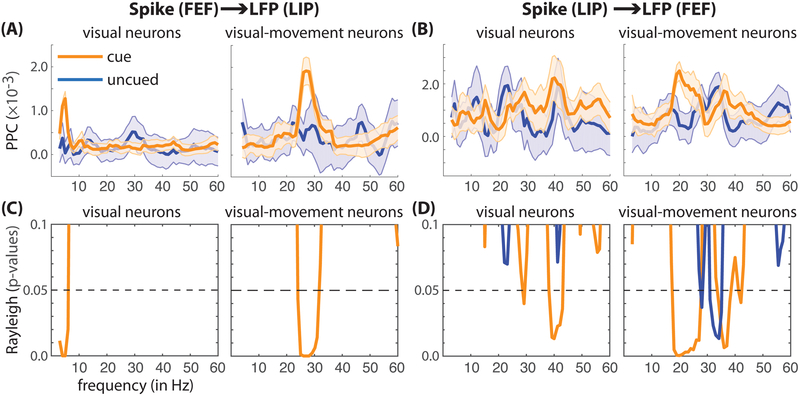
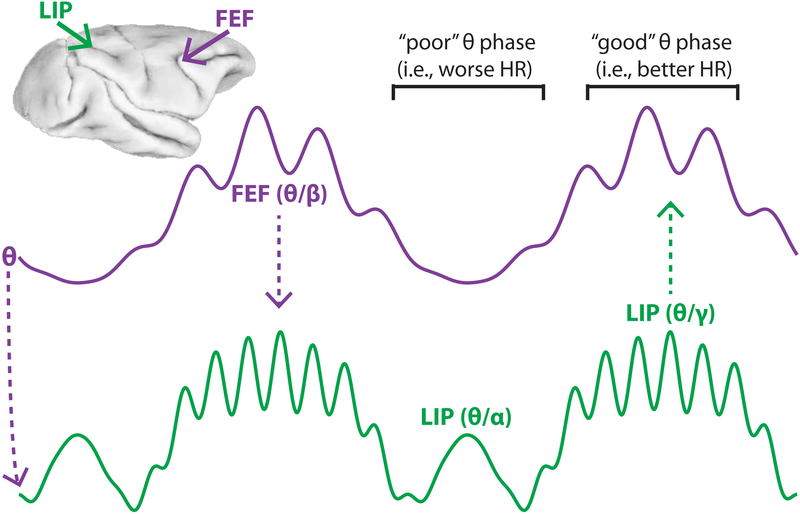
Classic studies of spatial attention assumed that its neural and behavioral effects were continuous over time. Recent behavioral studies have instead revealed that spatial attention leads to alternating periods of heightened or diminished perceptual sensitivity. Yet, the neural basis of these rhythmic fluctuations has remained largely unknown. We show that a dynamic interplay within the macaque frontoparietal network accounts for the rhythmic properties of spatial attention. Neural oscillations characterize functional interactions between the frontal eye fields (FEF) and the lateral intraparietal area (LIP), with theta phase (3-8 Hz) coordinating two rhythmically alternating states. The first is defined by FEF-dominated beta-band activity, associated with suppressed attentional shifts, and LIP-dominated gamma-band activity, associated with enhanced visual processing and better behavioral performance. The second is defined by LIP-specific alpha-band activity, associated with attenuated visual processing and worse behavioral performance. Our findings reveal how network-level interactions organize environmental sampling into rhythmic cycles.
Keywords: FEF; LIP; attention; frontoparietal network; oscillations; phase-dependent behavior; theta; vision.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
Comment in
-
Attention Cycles.Neuron. 2018 Aug 22;99(4):632-634. doi: 10.1016/j.neuron.2018.08.006. Neuron. 2018. PMID: 30138586
References
-
- Aru J, Aru J, Priesemann V, Wibral M, Lana L, Pipa G, Singer W, and Vicente R (2015). Untangling cross-frequency coupling in neuroscience. Current opinion in neurobiology 31, 51–61. - PubMed
-
- Barash S, Bracewell RM, Fogassi L, Gnadt JW, and Andersen RA (1991a). Saccade-related activity in the lateral intraparietal area. I. Temporal properties; comparison with area 7a. Journal of neurophysiology 66, 1095–1108. - PubMed
-
- Barash S, Bracewell RM, Fogassi L, Gnadt JW, and Andersen RA (1991). Saccade-related activity in the lateral intraparietal area. II. Spatial properties. Journal of neurophysiology 66, 1109–1124. - PubMed
-
- Bastos AM, Vezoli J, Bosman CA, Schoffelen JM, Oostenveld R, Dowdall JR, De Weerd P, Kennedy H, and Fries P (2015). Visual Areas Exert Feedforward and Feedback Influences through Distinct Frequency Channels. Neuron 85, 390–401. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
