

Severing enzymes amplify microtubule arrays through lattice GTP-tubulin incorporation
- PMID: 30139843
- PMCID: PMC6510489
- DOI: 10.1126/science.aau1504
Severing enzymes amplify microtubule arrays through lattice GTP-tubulin incorporation
Abstract
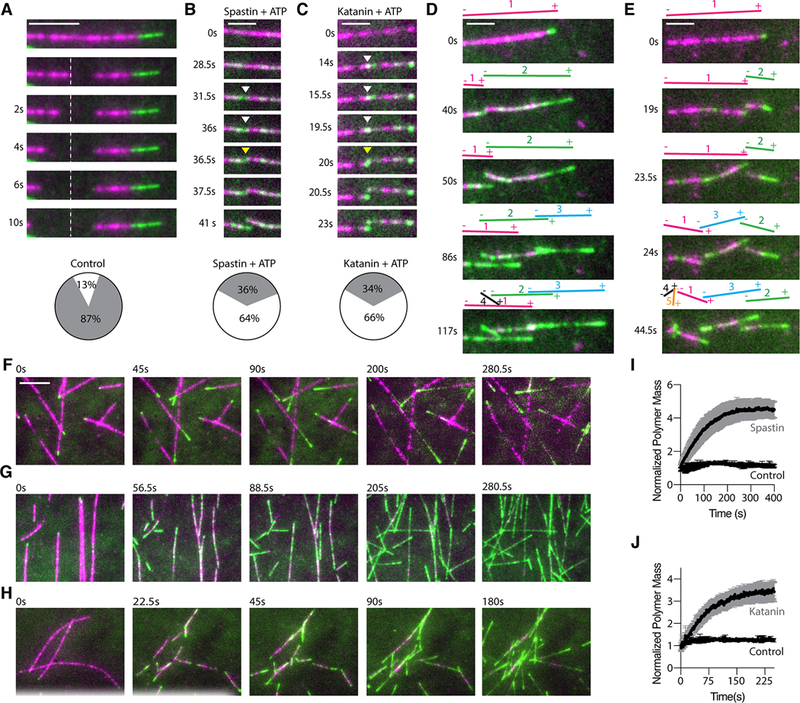
Spastin and katanin sever and destabilize microtubules. Paradoxically, despite their destructive activity they increase microtubule mass in vivo. We combined single-molecule total internal reflection fluorescence microscopy and electron microscopy to show that the elemental step in microtubule severing is the generation of nanoscale damage throughout the microtubule by active extraction of tubulin heterodimers. These damage sites are repaired spontaneously by guanosine triphosphate (GTP)-tubulin incorporation, which rejuvenates and stabilizes the microtubule shaft. Consequently, spastin and katanin increase microtubule rescue rates. Furthermore, newly severed ends emerge with a high density of GTP-tubulin that protects them against depolymerization. The stabilization of the newly severed plus ends and the higher rescue frequency synergize to amplify microtubule number and mass. Thus, severing enzymes regulate microtubule architecture and dynamics by promoting GTP-tubulin incorporation within the microtubule shaft.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures






Comment in
-
Strengthening Microtubules by Cuts that Heal.Dev Cell. 2018 Nov 19;47(4):400-401. doi: 10.1016/j.devcel.2018.11.002. Dev Cell. 2018. PMID: 30458133
-
What Doesn't Kill You Makes You Stronger.Dev Cell. 2018 Nov 19;47(4):402-403. doi: 10.1016/j.devcel.2018.11.003. Dev Cell. 2018. PMID: 30458134
References
-
- Trotta N, Orso G, Rossetto MG, Daga A, Broadie K, The hereditary spastic paraplegia gene, spastin, regulates microtubule stability to modulate synaptic structure and function. Current Biology 14, 1135–1147 (2004). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
