

Polycomb complexes associate with enhancers and promote oncogenic transcriptional programs in cancer through multiple mechanisms
- PMID: 30139998
- PMCID: PMC6107513
- DOI: 10.1038/s41467-018-05728-x
Polycomb complexes associate with enhancers and promote oncogenic transcriptional programs in cancer through multiple mechanisms
Abstract
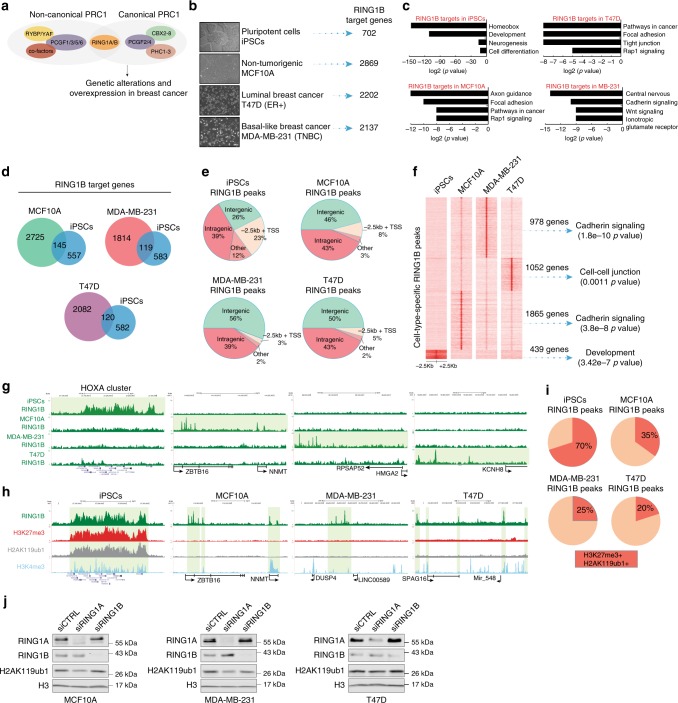
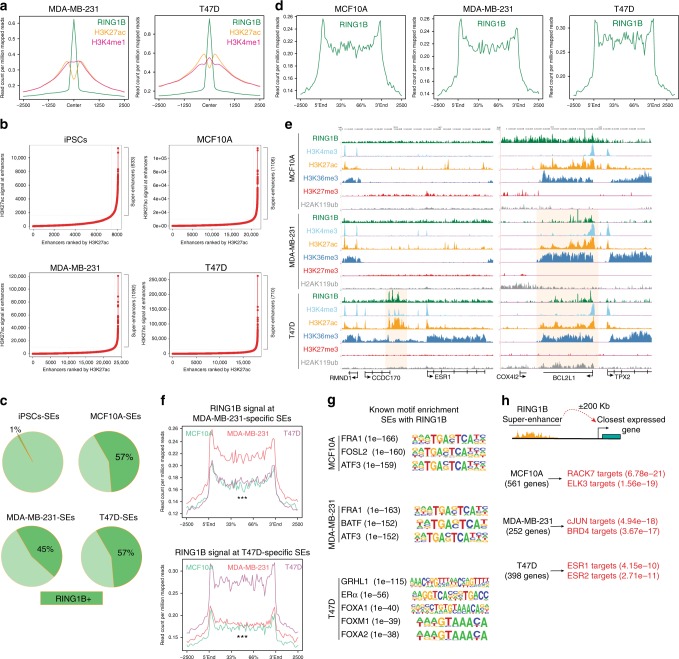
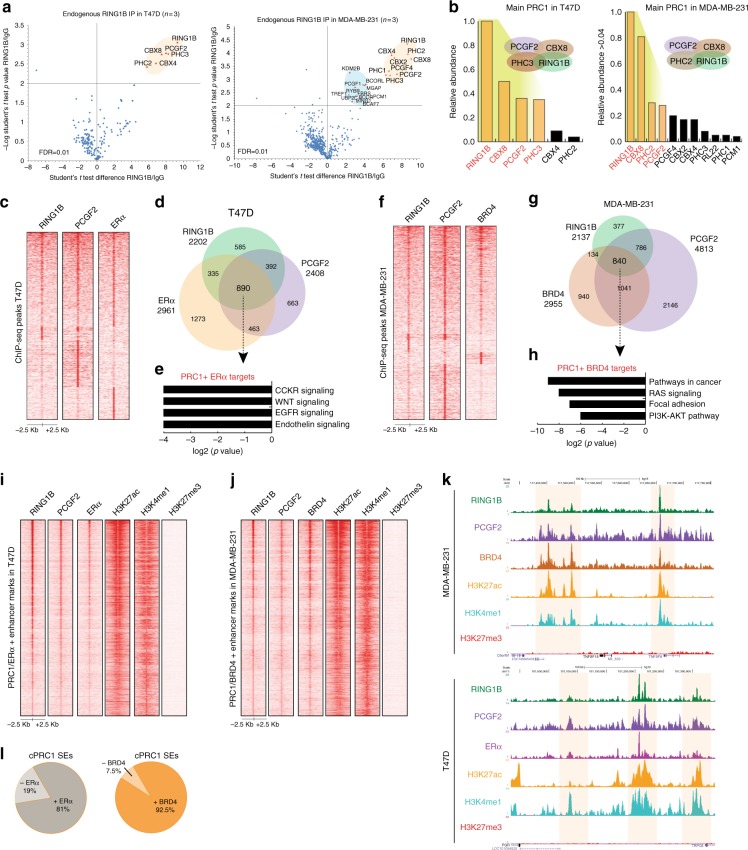
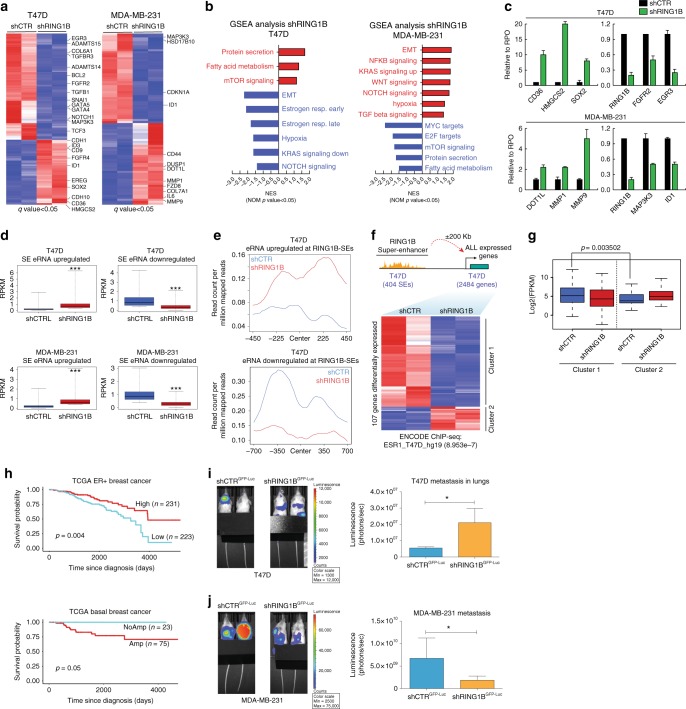
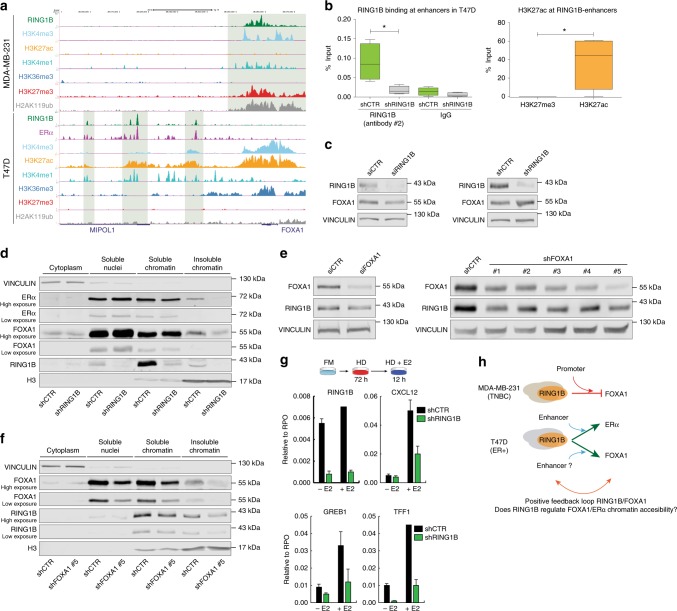
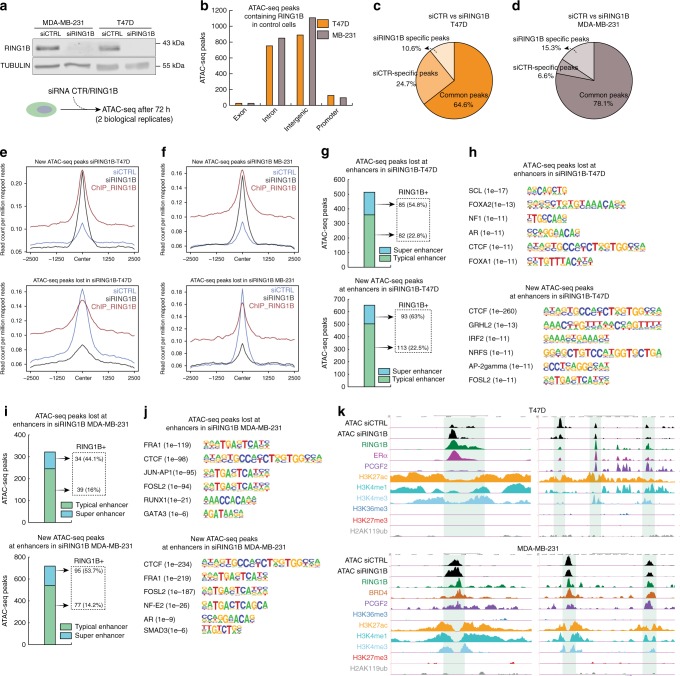
Polycomb repressive complex 1 (PRC1) plays essential roles in cell fate decisions and development. However, its role in cancer is less well understood. Here, we show that RNF2, encoding RING1B, and canonical PRC1 (cPRC1) genes are overexpressed in breast cancer. We find that cPRC1 complexes functionally associate with ERα and its pioneer factor FOXA1 in ER+ breast cancer cells, and with BRD4 in triple-negative breast cancer cells (TNBC). While cPRC1 still exerts its repressive function, it is also recruited to oncogenic active enhancers. RING1B regulates enhancer activity and gene transcription not only by promoting the expression of oncogenes but also by regulating chromatin accessibility. Functionally, RING1B plays a divergent role in ER+ and TNBC metastasis. Finally, we show that concomitant recruitment of RING1B to active enhancers occurs across multiple cancers, highlighting an under-explored function of cPRC1 in regulating oncogenic transcriptional programs in cancer.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Comet I, Riising EM, Leblanc B, Helin K. Maintaining cell identity: PRC2-mediated regulation of transcription and cancer. Nat. Publ. Group. 2016;16:803–810. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
