

Structure and mechanism of cancer-associated N-acetylglucosaminyltransferase-V
- PMID: 30140003
- PMCID: PMC6107550
- DOI: 10.1038/s41467-018-05931-w
Structure and mechanism of cancer-associated N-acetylglucosaminyltransferase-V
Abstract
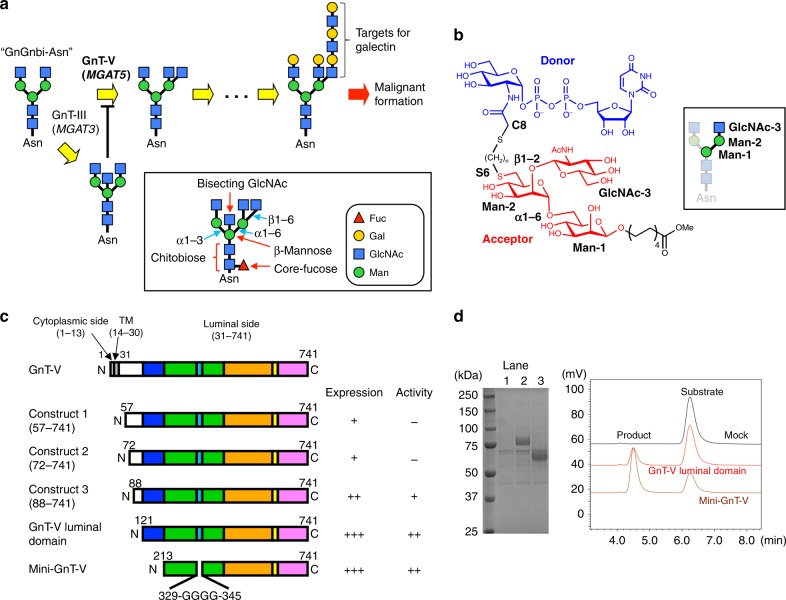
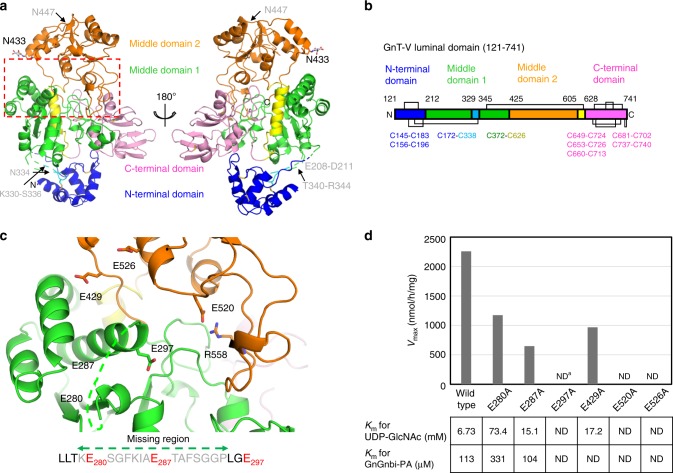
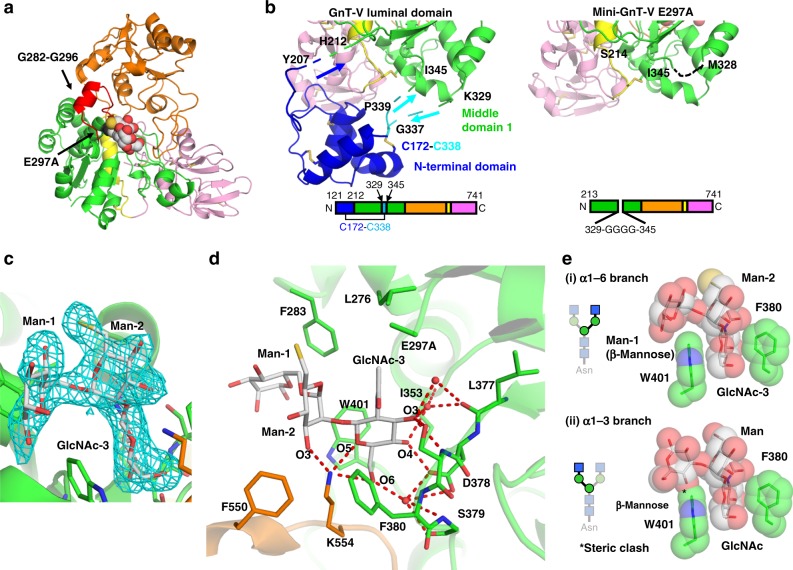
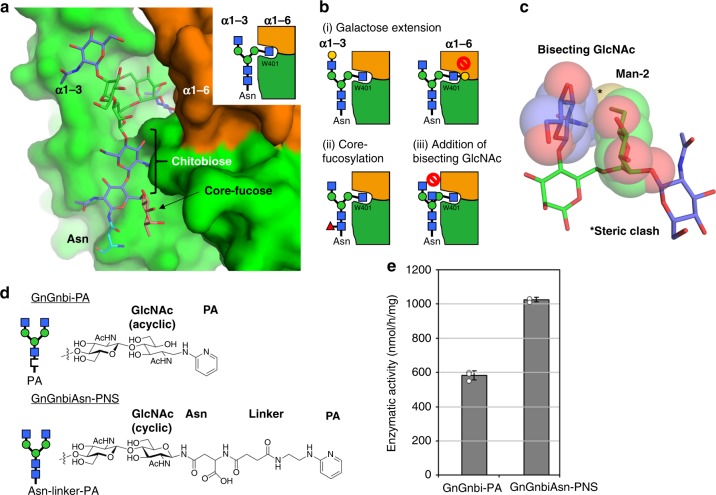
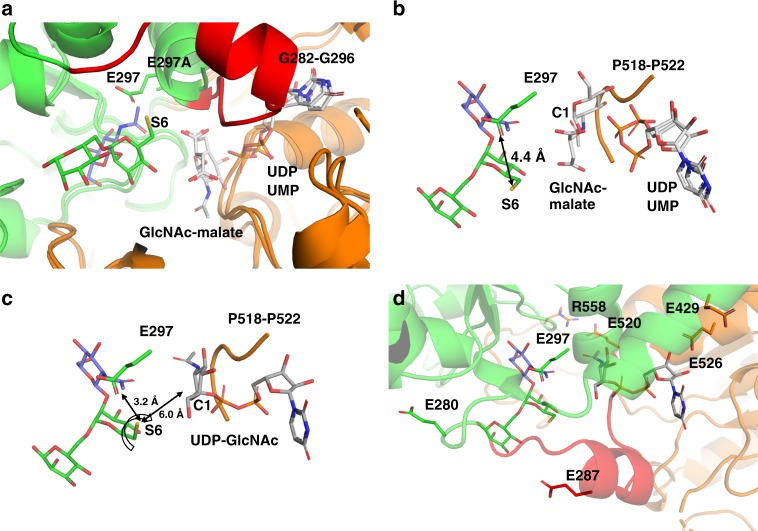
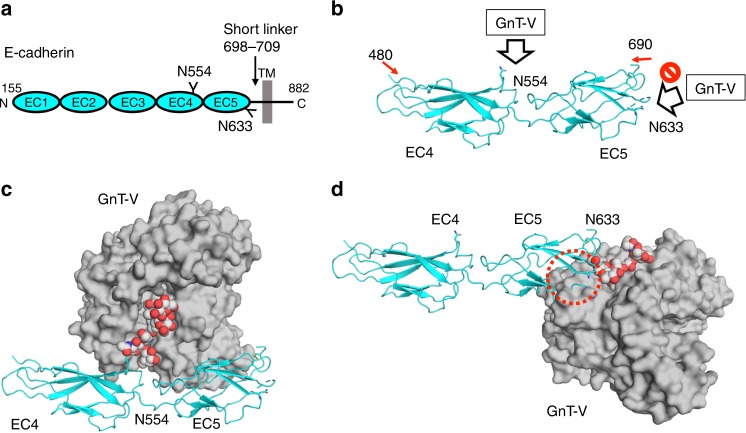
N-acetylglucosaminyltransferase-V (GnT-V) alters the structure of specific N-glycans by modifying α1-6-linked mannose with a β1-6-linked N-acetylglucosamine branch. β1-6 branch formation on cell surface receptors accelerates cancer metastasis, making GnT-V a promising target for drug development. However, the molecular basis of GnT-V's catalytic mechanism and substrate specificity are not fully understood. Here, we report crystal structures of human GnT-V luminal domain with a substrate analog. GnT-V luminal domain is composed of a GT-B fold and two accessary domains. Interestingly, two aromatic rings sandwich the α1-6 branch of the acceptor N-glycan and restrain the global conformation, partly explaining the fine branch specificity of GnT-V. In addition, interaction of the substrate N-glycoprotein with GnT-V likely contributes to protein-selective and site-specific glycan modification. In summary, the acceptor-GnT-V complex structure suggests a catalytic mechanism, explains the previously observed inhibition of GnT-V by branching enzyme GnT-III, and provides a basis for the rational design of drugs targeting N-glycan branching.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Substrate Preference and Interplay of Fucosyltransferase 8 and N-Acetylglucosaminyltransferases.J Am Chem Soc. 2017 Jul 19;139(28):9431-9434. doi: 10.1021/jacs.7b03729. Epub 2017 Jul 11. J Am Chem Soc. 2017. PMID: 28678517
-
Recognition of glycan and protein substrates by N-acetylglucosaminyltransferase-V.Biochim Biophys Acta Gen Subj. 2020 Dec;1864(12):129726. doi: 10.1016/j.bbagen.2020.129726. Epub 2020 Sep 2. Biochim Biophys Acta Gen Subj. 2020. PMID: 32890705
-
N-acetylglucosaminyltransferase-V requires a specific noncatalytic luminal domain for its activity toward glycoprotein substrates.J Biol Chem. 2022 Mar;298(3):101666. doi: 10.1016/j.jbc.2022.101666. Epub 2022 Jan 30. J Biol Chem. 2022. PMID: 35104505 Free PMC article.
-
Structure and function of N-acetylglucosaminyltransferase V (GnT-V).Biochim Biophys Acta Gen Subj. 2024 Nov;1868(11):130709. doi: 10.1016/j.bbagen.2024.130709. Epub 2024 Sep 2. Biochim Biophys Acta Gen Subj. 2024. PMID: 39233219 Review.
-
Regulation of human GnT-IV family activity by the lectin domain.Carbohydr Res. 2024 Nov;545:109285. doi: 10.1016/j.carres.2024.109285. Epub 2024 Oct 2. Carbohydr Res. 2024. PMID: 39369636 Review.
Cited by
-
Global mapping of glycosylation pathways in human-derived cells.Dev Cell. 2021 Apr 19;56(8):1195-1209.e7. doi: 10.1016/j.devcel.2021.02.023. Epub 2021 Mar 16. Dev Cell. 2021. PMID: 33730547 Free PMC article.
-
Cryostorage of unstable N-acetylglucosaminyltransferase-V by synthetic zwitterions.RSC Adv. 2022 Apr 14;12(19):11628-11631. doi: 10.1039/d2ra01575g. eCollection 2022 Apr 13. RSC Adv. 2022. PMID: 35481097 Free PMC article.
-
The Roles of Glycans in Bladder Cancer.Front Oncol. 2020 Jun 12;10:957. doi: 10.3389/fonc.2020.00957. eCollection 2020. Front Oncol. 2020. PMID: 32596162 Free PMC article. Review.
-
The Association between Urine N-Glycome and Prognosis after Initial Therapy for Primary Prostate Cancer.Biomedicines. 2024 May 8;12(5):1039. doi: 10.3390/biomedicines12051039. Biomedicines. 2024. PMID: 38791001 Free PMC article.
-
Structural glycobiology - from enzymes to organelles.Biochem Soc Trans. 2025 Jan 31;53(1):83-100. doi: 10.1042/BST20241119. Biochem Soc Trans. 2025. PMID: 39889286 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
