

The tuberous sclerosis complex subunit TBC1D7 is stabilized by Akt phosphorylation-mediated 14-3-3 binding
- PMID: 30143532
- PMCID: PMC6200923
- DOI: 10.1074/jbc.RA118.003525
The tuberous sclerosis complex subunit TBC1D7 is stabilized by Akt phosphorylation-mediated 14-3-3 binding
Abstract
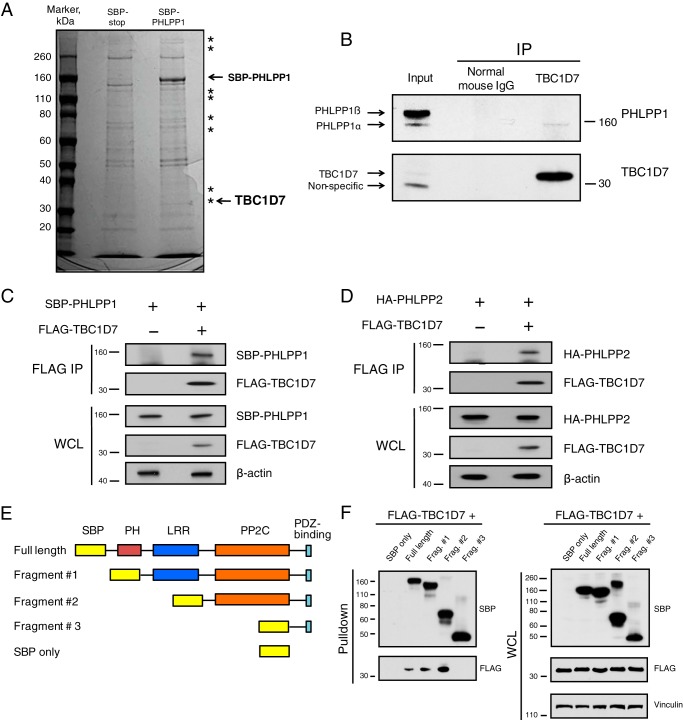
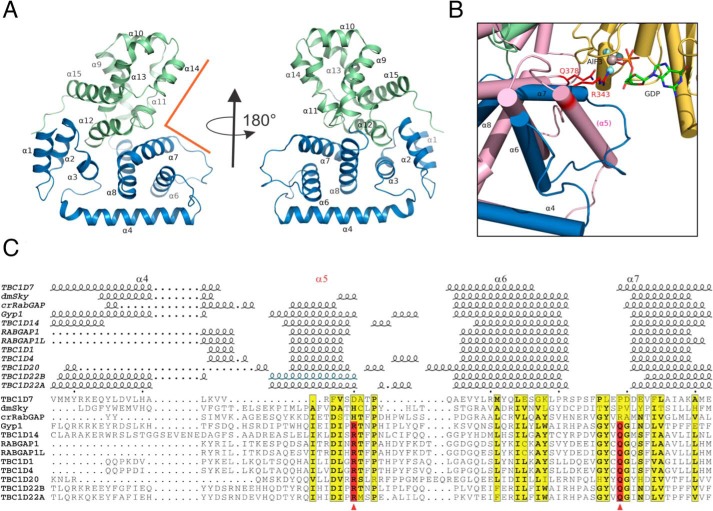
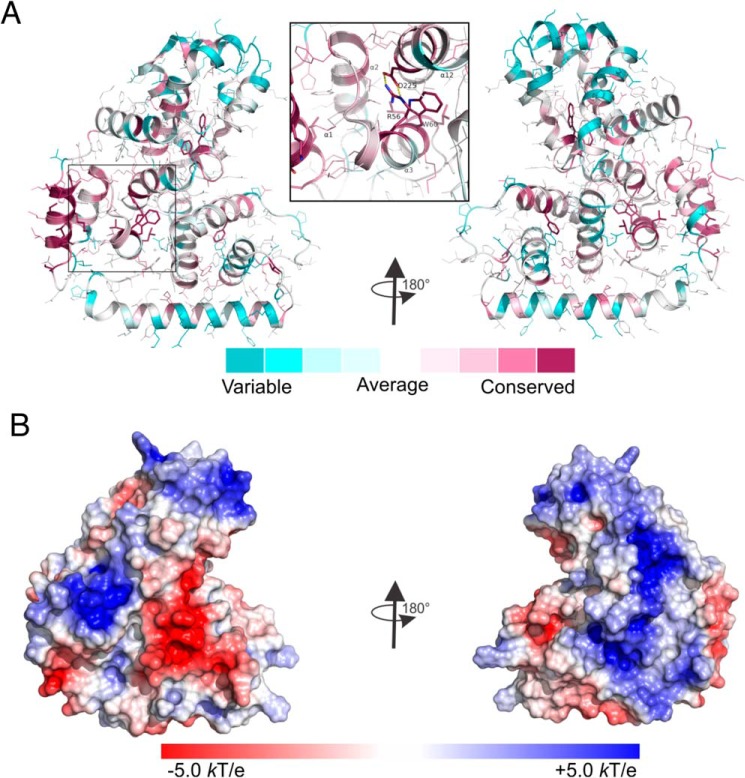
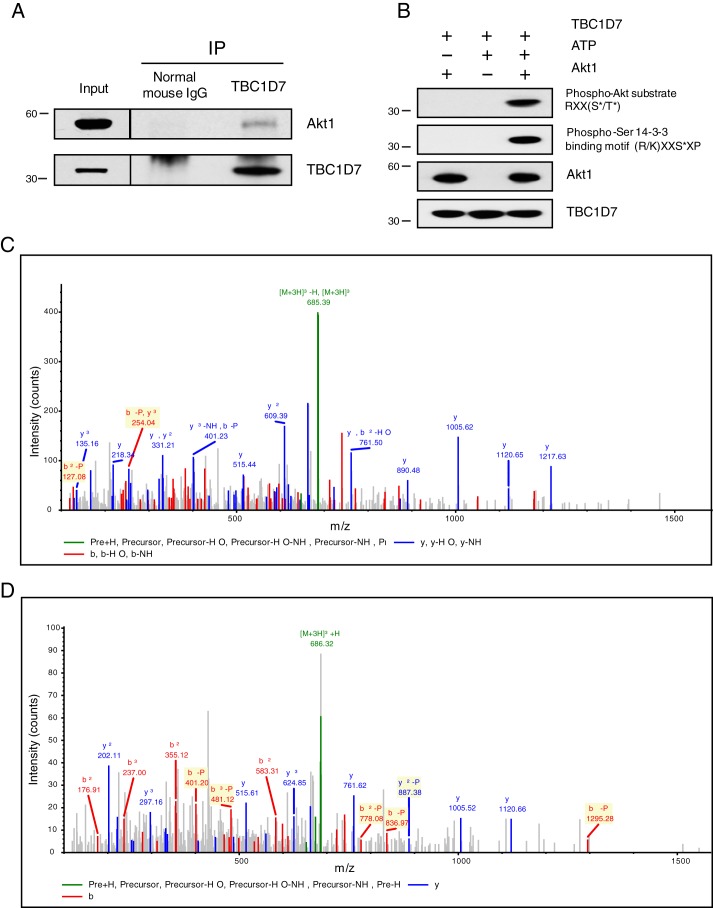
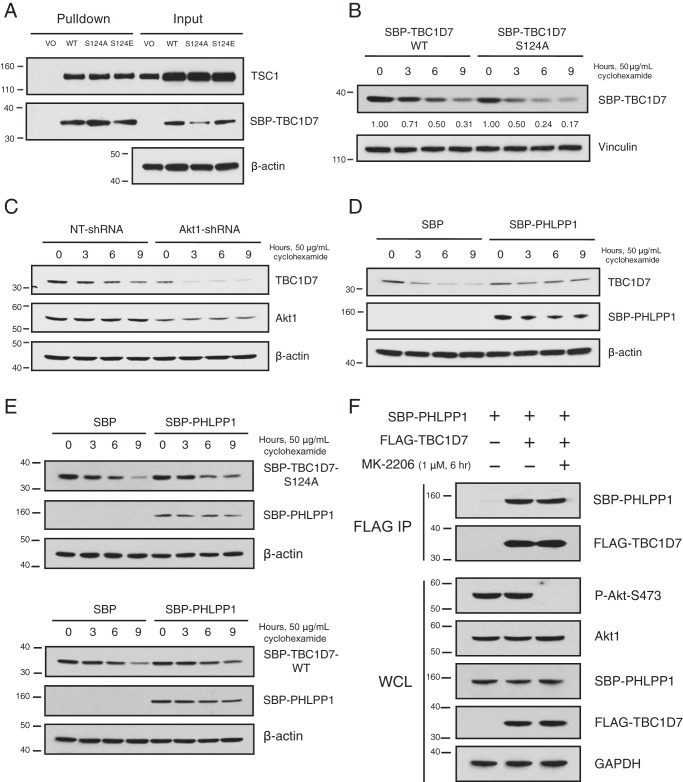
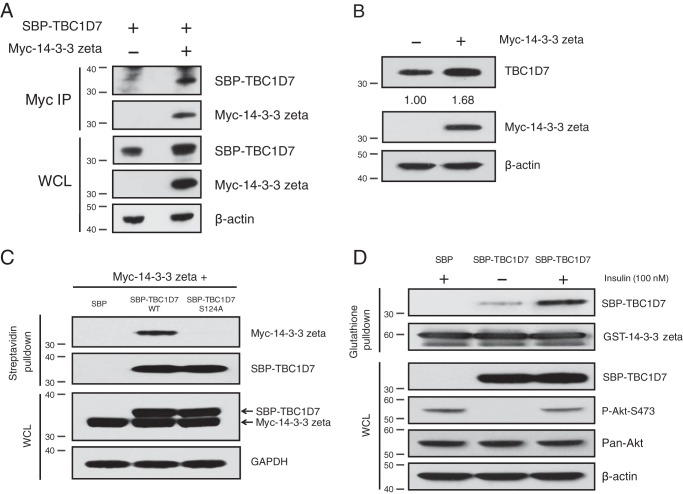
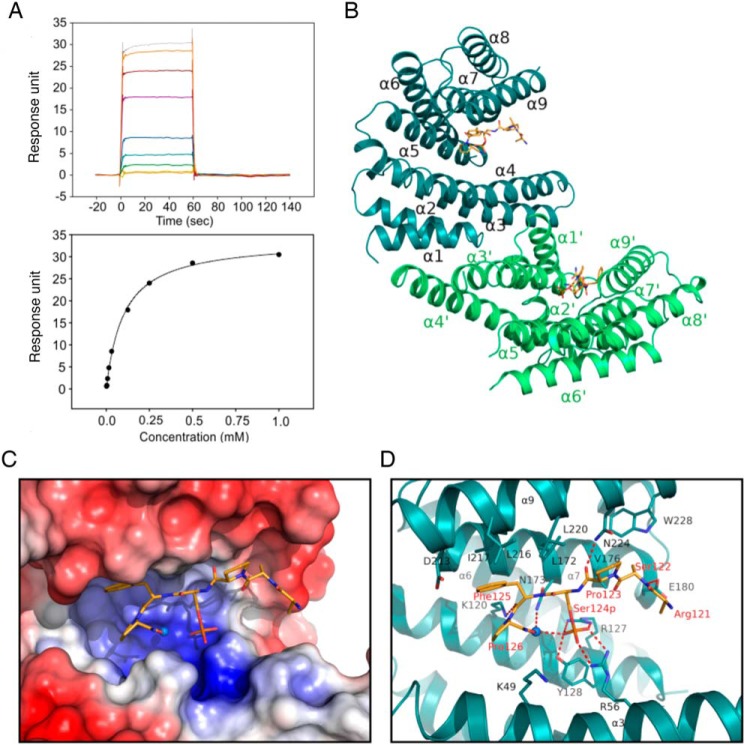
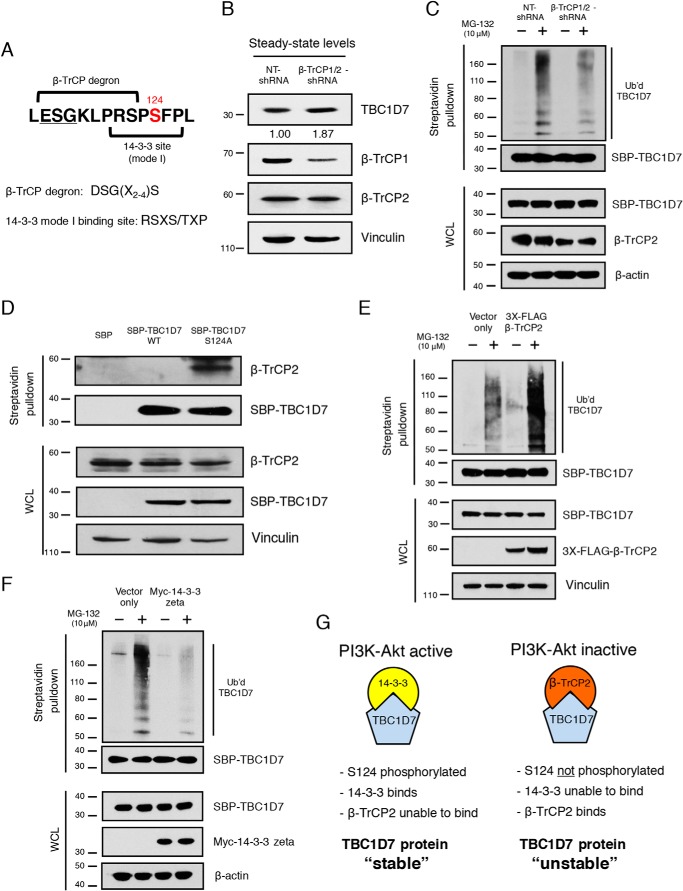
The tuberous sclerosis complex (TSC) is a negative regulator of mTOR complex 1, a signaling node promoting cellular growth in response to various nutrients and growth factors. However, several regulators in TSC signaling still await discovery and characterization. Using pulldown and MS approaches, here we identified the TSC complex member, TBC1 domain family member 7 (TBC1D7), as a binding partner for PH domain and leucine-rich repeat protein phosphatase 1 (PHLPP1), a negative regulator of Akt kinase signaling. Most TBC domain-containing proteins function as Rab GTPase-activating proteins (RabGAPs), but the crystal structure of TBC1D7 revealed that it lacks residues critical for RabGAP activity. Sequence analysis identified a putative site for both Akt-mediated phosphorylation and 14-3-3 binding at Ser-124, and we found that Akt phosphorylates TBC1D7 at Ser-124. However, this phosphorylation had no effect on the binding of TBC1D7 to TSC1, but stabilized TBC1D7. Moreover, 14-3-3 protein both bound and stabilized TBC1D7 in a growth factor-dependent manner, and a phospho-deficient substitution, S124A, prevented this interaction. The crystal structure of 14-3-3ζ in complex with a phospho-Ser-124 TBC1D7 peptide confirmed the direct interaction between 14-3-3 and TBC1D7. The sequence immediately upstream of Ser-124 aligned with a canonical β-TrCP degron, and we found that the E3 ubiquitin ligase β-TrCP2 ubiquitinates TBC1D7 and decreases its stability. Our findings reveal that Akt activity determines the phosphorylation status of TBC1D7 at the phospho-switch Ser-124, which governs binding to either 14-3-3 or β-TrCP2, resulting in increased or decreased stability of TBC1D7, respectively.
Keywords: 14-3-3 protein; Akt PKB; E3 ubiquitin ligase; mTORC1; phospho-switch; phosphorylation; protein stability; tuberous sclerosis complex (TSC); ubiquitylation (ubiquitination).
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
