

Lactate transport facilitates neurite outgrowth
- PMID: 30143583
- PMCID: PMC6167502
- DOI: 10.1042/BSR20180157
Lactate transport facilitates neurite outgrowth
Abstract
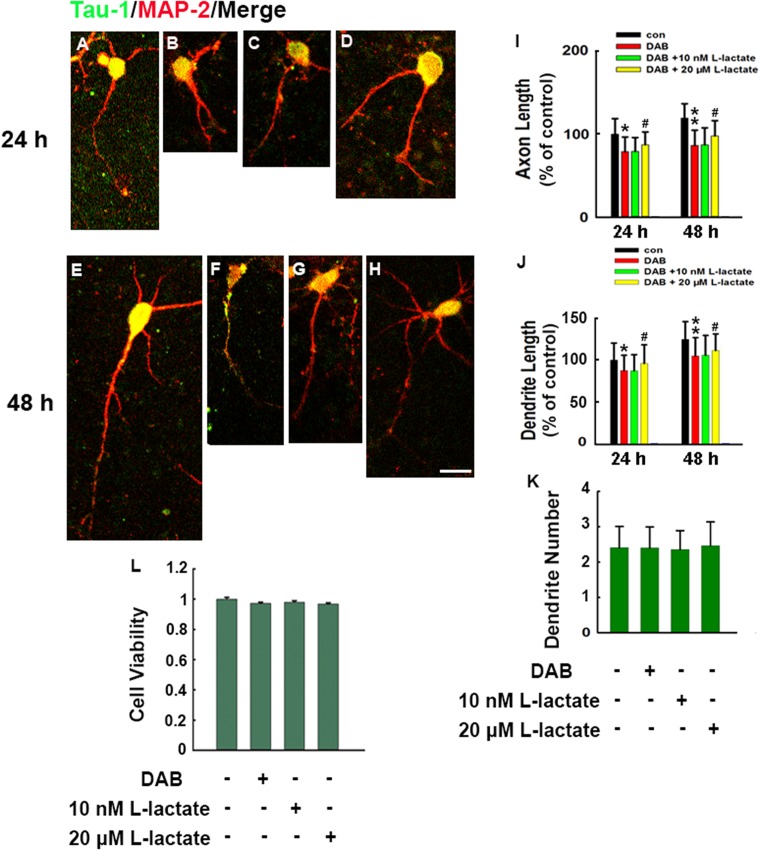
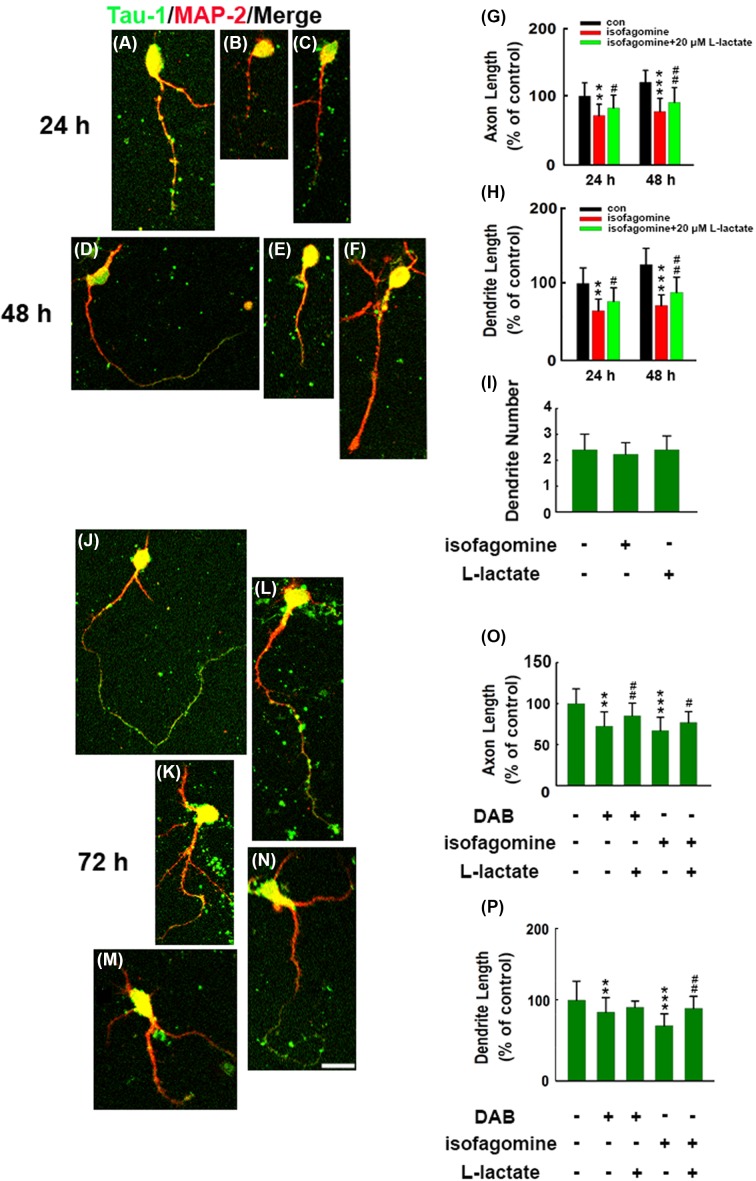
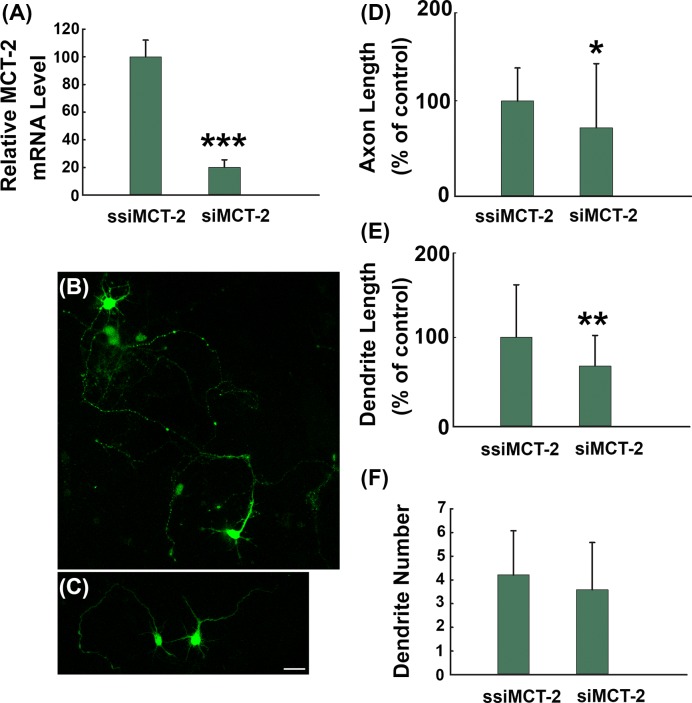
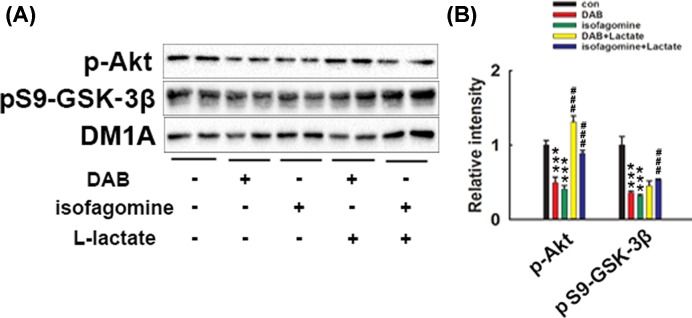
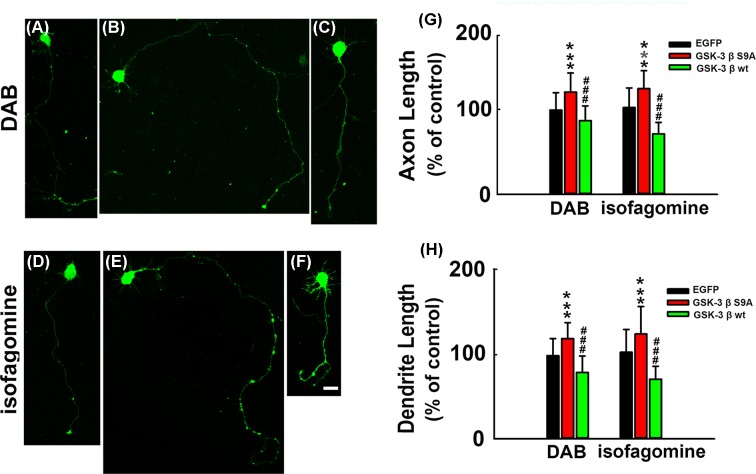
How glia affect neurite outgrowth during neural development has not been well elucidated. In the present study, we found that disruption of lactate production using 1,4-dideoxy-1,4-imino-D-arabinitol (DAB) and isofagomine significantly interfered with neurite outgrowth and that exogenous application of L-lactate rescued neurite growth failure. Monocarboxylate transporter-2-knockout, which blocked the lactate shuttle in neurons, showed a remarkable decrease in the length of axons and dendrites. We further demonstrated that Akt activity was decreased while glycogen synthase kinase 3β (GSK3β) activity was increased after astrocytic glycogen phosphorylase blockade. Additionally, GSK3βSer9 mutation reversed neurite growth failure caused by DAB and isofagomine. Our results suggested that lactate transportation played a critical role in neural development and disruption of the lactate shuttle in quiescent condition also affected neurite outgrowth in the central nervous system.
Keywords: Akt; Glycogen synthase kinase 3β (GSK-3β); L-lactate; Monocarboxylate transporters-2 (MCT-2); Neurite outgrowth.
© 2018 The Author(s).
Conflict of interest statement
The authors declare that there are no competing interests associated with the manuscript.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases