

Differential Induction of IFN-α and Modulation of CD112 and CD54 Expression Govern the Magnitude of NK Cell IFN-γ Response to Influenza A Viruses
- PMID: 30143589
- PMCID: PMC6143432
- DOI: 10.4049/jimmunol.1800161
Differential Induction of IFN-α and Modulation of CD112 and CD54 Expression Govern the Magnitude of NK Cell IFN-γ Response to Influenza A Viruses
Abstract
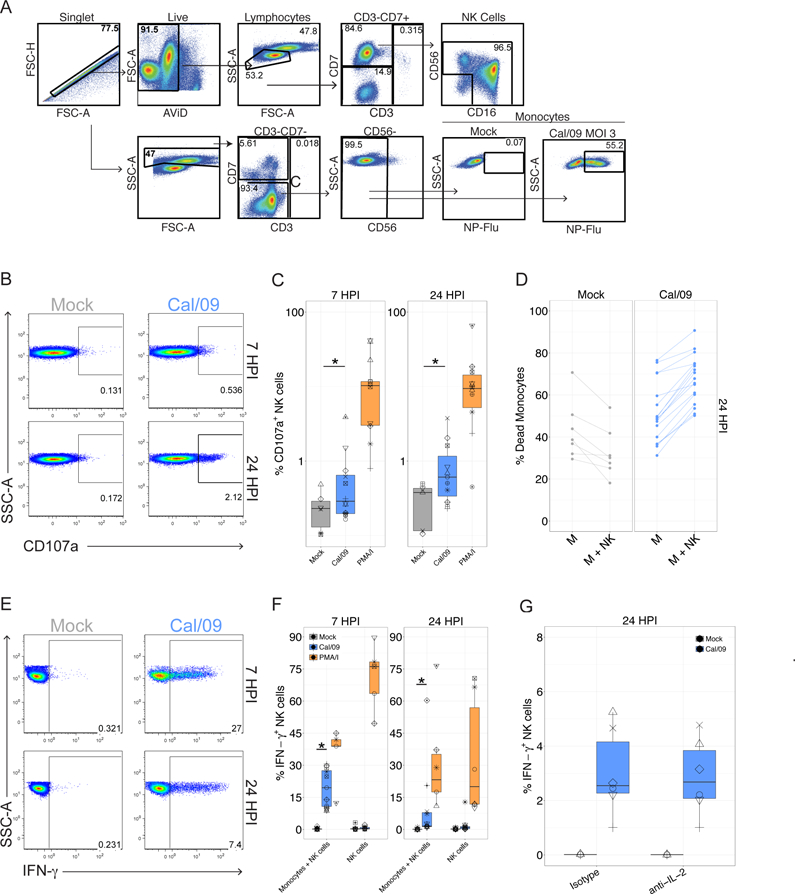
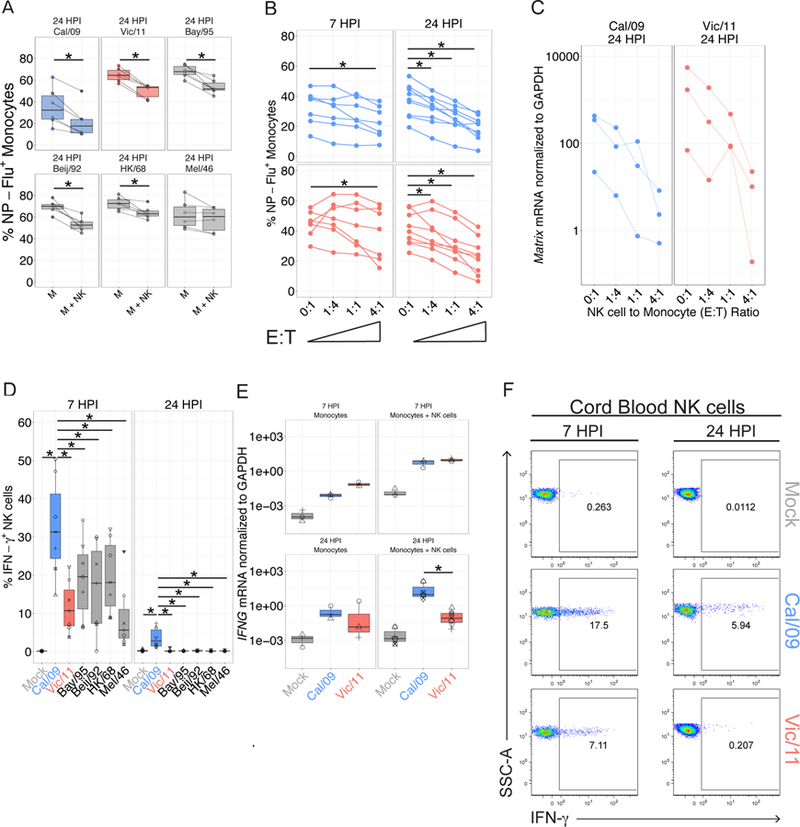
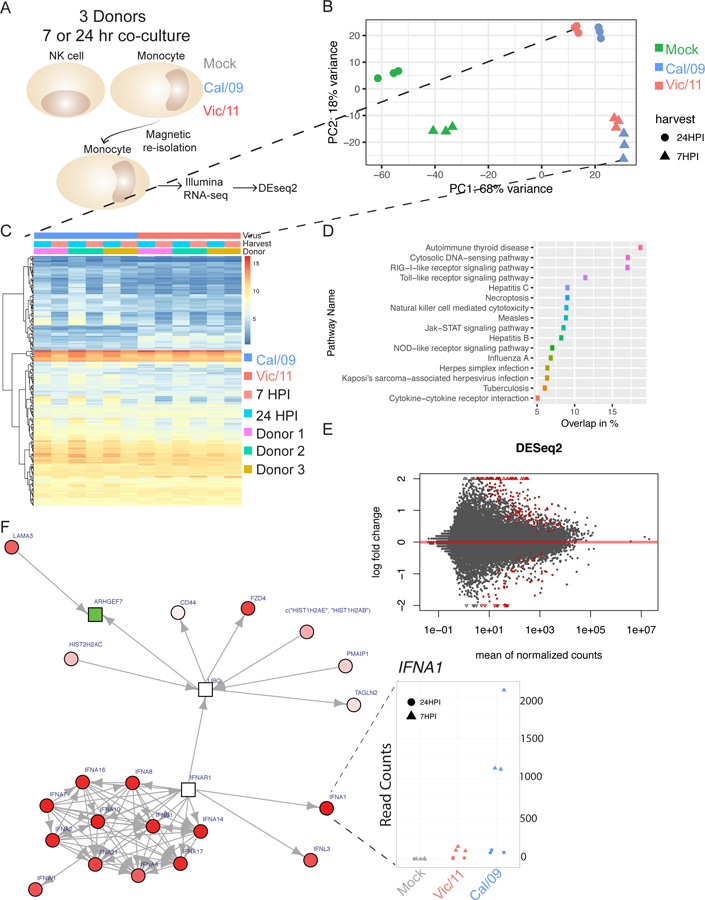
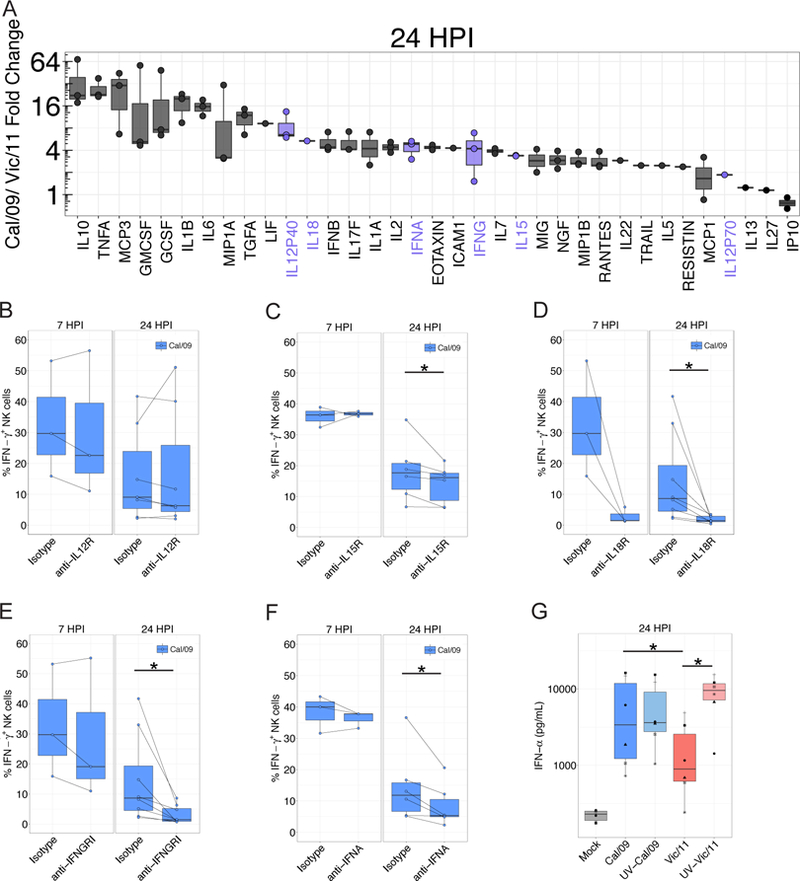
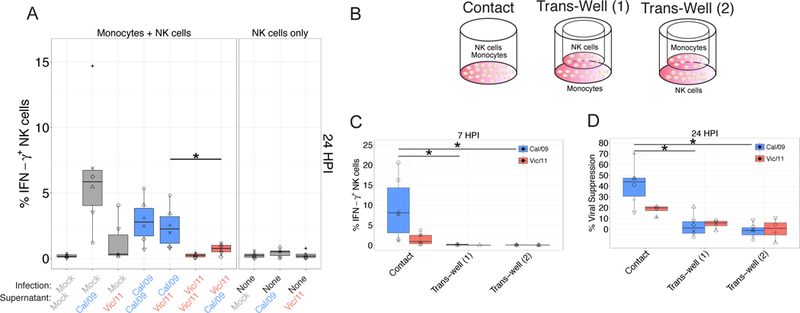
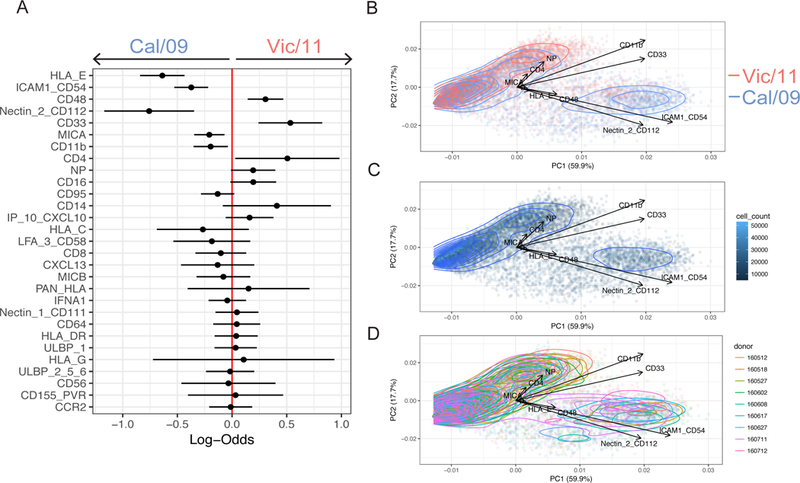
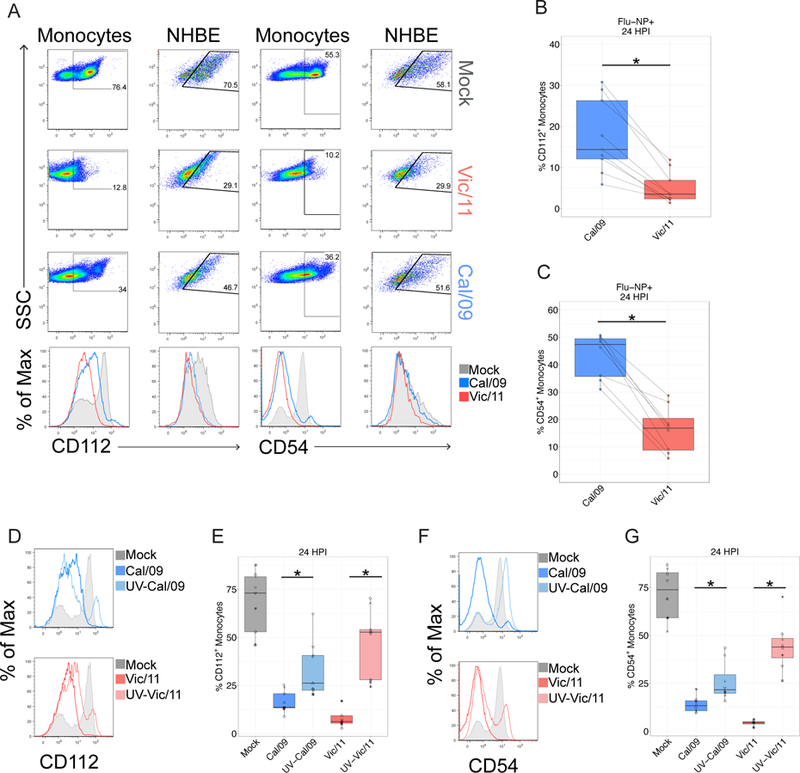
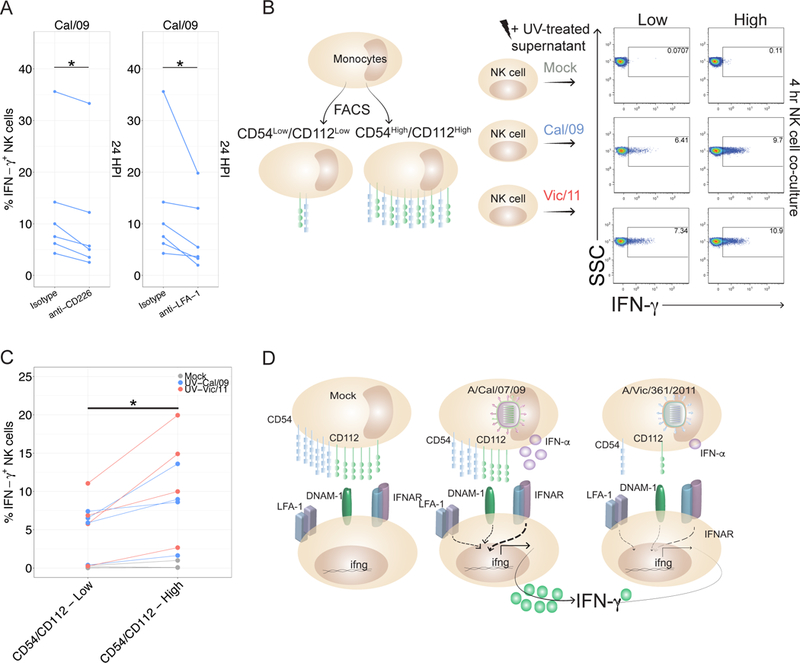
In human and murine studies, IFN-γ is a critical mediator immunity to influenza. IFN-γ production is critical for viral clearance and the development of adaptive immune responses, yet excessive production of IFN-γ and other cytokines as part of a cytokine storm is associated with poor outcomes of influenza infection in humans. As NK cells are the main population of lung innate immune cells capable of producing IFN-γ early in infection, we set out to identify the drivers of the human NK cell IFN-γ response to influenza A viruses. We found that influenza triggers NK cells to secrete IFN-γ in the absence of T cells and in a manner dependent upon signaling from both cytokines and receptor-ligand interactions. Further, we discovered that the pandemic A/California/07/2009 (H1N1) strain elicits a seven-fold greater IFN-γ response than other strains tested, including a seasonal A/Victoria/361/2011 (H3N2) strain. These differential responses were independent of memory NK cells. Instead, we discovered that the A/Victoria/361/2011 influenza strain suppresses the NK cell IFN-γ response by downregulating NK-activating ligands CD112 and CD54 and by repressing the type I IFN response in a viral replication-dependent manner. In contrast, the A/California/07/2009 strain fails to repress the type I IFN response or to downregulate CD54 and CD112 to the same extent, which leads to the enhanced NK cell IFN-γ response. Our results indicate that influenza implements a strain-specific mechanism governing NK cell production of IFN-γ and identifies a previously unrecognized influenza innate immune evasion strategy.
Copyright © 2018 by The American Association of Immunologists, Inc.
Conflict of interest statement
The authors have declared that no conflict of interest exists.
Figures
References
-
- Ge MQ, Ho AWS, Tang Y, Wong KHS, Chua BYL, Gasser S, and Kemeny DM. 2012. NK cells regulate CD8+ T cell priming and dendritic cell migration during influenza A infection by IFN-γ and perforin-dependent mechanisms. J. Immunol 189: 2099–2109. - PubMed
-
- Stein-Streilein J, Bennett M, Mann D, and Kumar V. 1983. Natural killer cells in mouse lung: surface phenotype, target preference, and response to local influenza virus infection. J. Immunol 131: 2699–2704. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
