

Structure of a Novel Dimeric SET Domain Methyltransferase that Regulates Cell Motility
- PMID: 30148980
- PMCID: PMC7141177
- DOI: 10.1016/j.jmb.2018.08.017
Structure of a Novel Dimeric SET Domain Methyltransferase that Regulates Cell Motility
Abstract
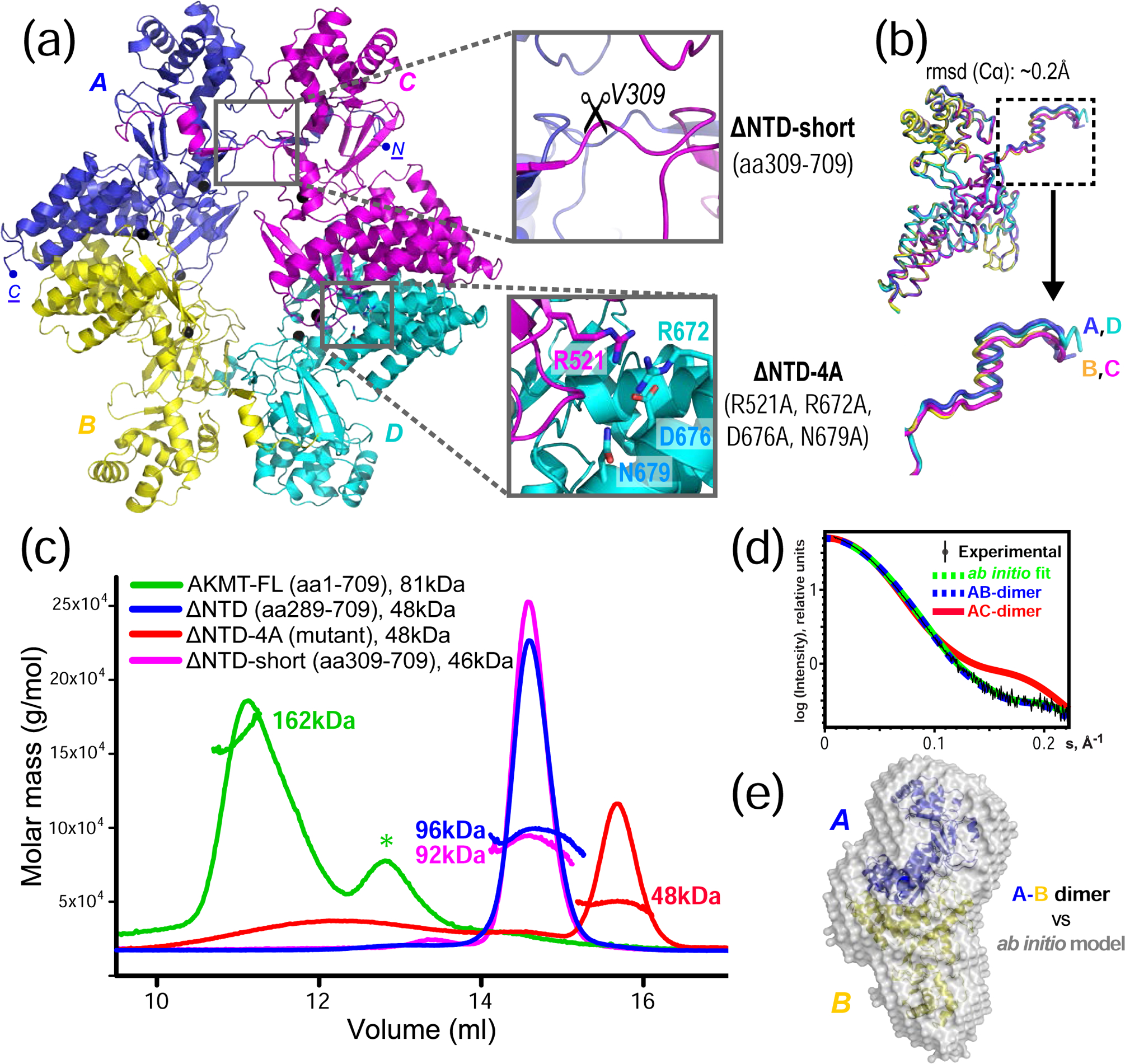
Lysine methyltransferases (KMTs) were initially associated with transcriptional control through their methylation of histones and other nuclear proteins, but have since been found to regulate many other cellular activities. The apical complex lysine (K) methyltransferase (AKMT) of the human parasite Toxoplasma gondii was recently shown to play a critical role in regulating cellular motility. Here we report a 2.1-Å resolution crystal structure of the conserved and functional C-terminal portion (aa289-709) of T. gondii AKMT. AKMT dimerizes via a unique intermolecular interface mediated by the C-terminal tetratricopeptide repeat-like domain together with a specific zinc-binding motif that is absent from all other KMTs. Disruption of AKMT dimerization impaired both its enzyme activity and parasite egress from infected host cells in vivo. Structural comparisons reveal that AKMT is related to the KMTs in the SMYD family, with, however, a number of distinct structural features in addition to the unusual dimerization interface. These features are conserved among the apicomplexan parasites and their free-living relatives, but not found in any known KMTs in animals. AKMT therefore is the founding member of a new subclass of KMT that has important implications for the evolution of the apicomplexans.
Keywords: AKMT; egress; lysine methylation; parasite.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Conflict of interest
The authors declare that they have no conflict of interest.
Figures







References
-
- Santos-Rosa H, et al. , Active genes are tri-methylated at K4 of histone H3. Nature, 2002. 419(6905): p. 407–11. - PubMed
-
- Huang J and Berger SL, The emerging field of dynamic lysine methylation of non-histone proteins. Curr Opin Genet Dev, 2008. 18(2): p. 152–8. - PubMed
-
- Yang XD, Lamb A, and Chen LF, Methylation, a new epigenetic mark for protein stability. Epigenetics, 2009. 4(7): p. 429–33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
