

Human iPS derived progenitors bioengineered into liver organoids using an inverted colloidal crystal poly (ethylene glycol) scaffold
- PMID: 30149262
- PMCID: PMC6131727
- DOI: 10.1016/j.biomaterials.2018.07.043
Human iPS derived progenitors bioengineered into liver organoids using an inverted colloidal crystal poly (ethylene glycol) scaffold
Abstract
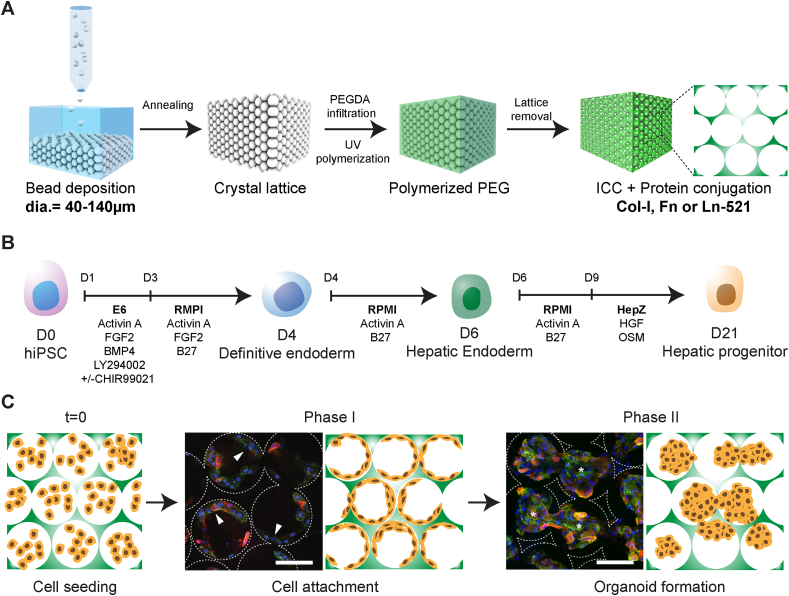
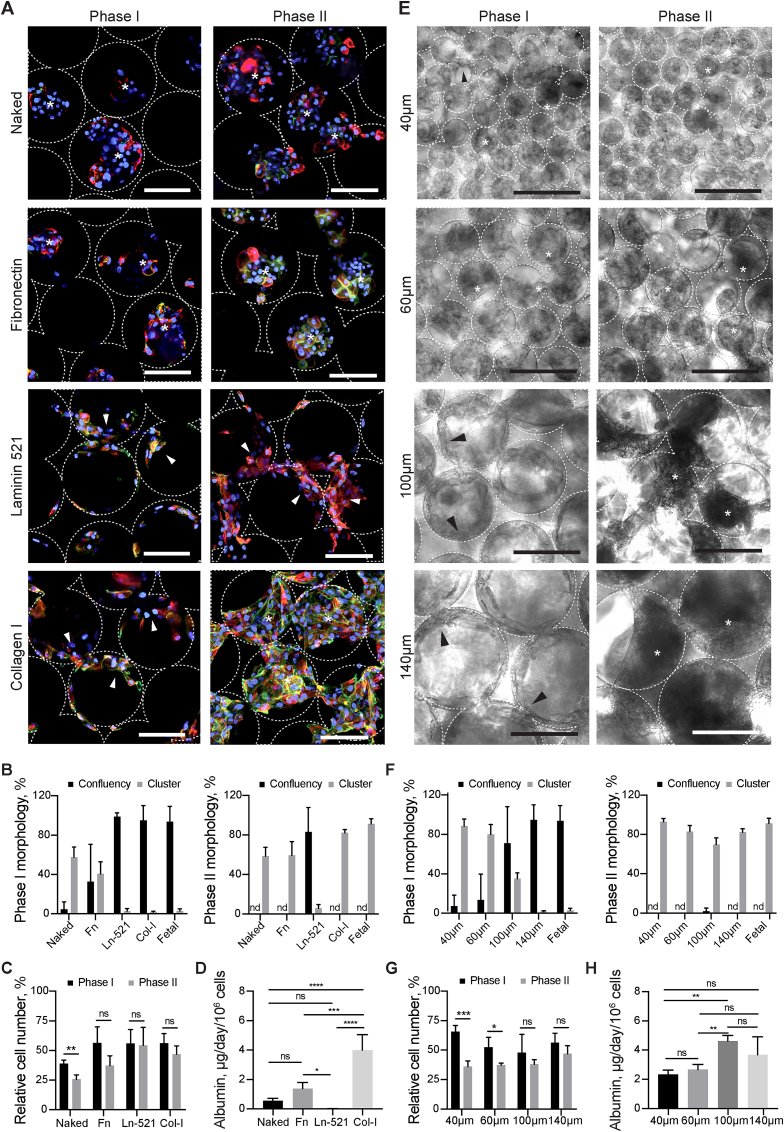
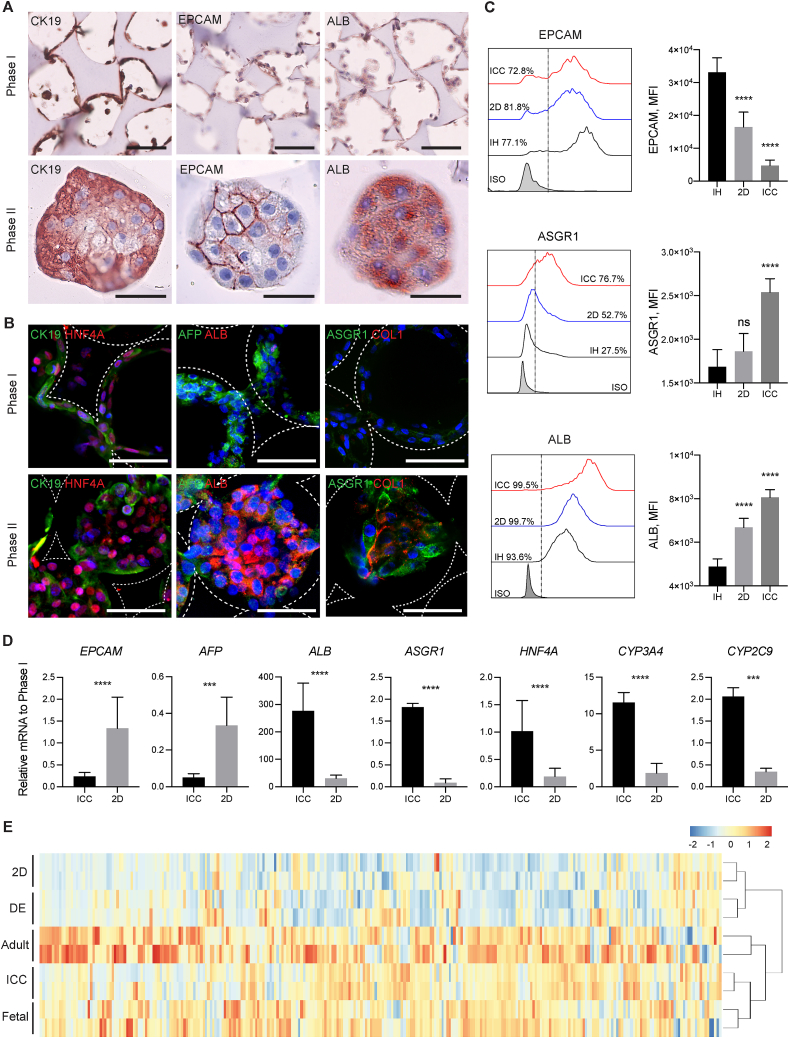
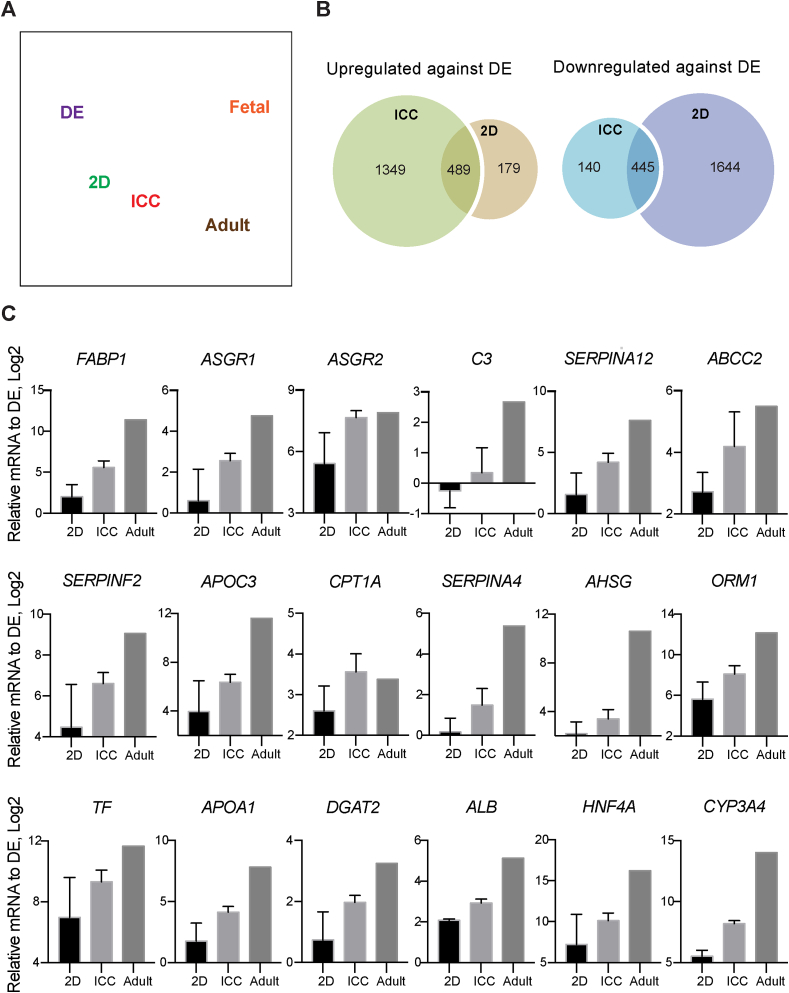
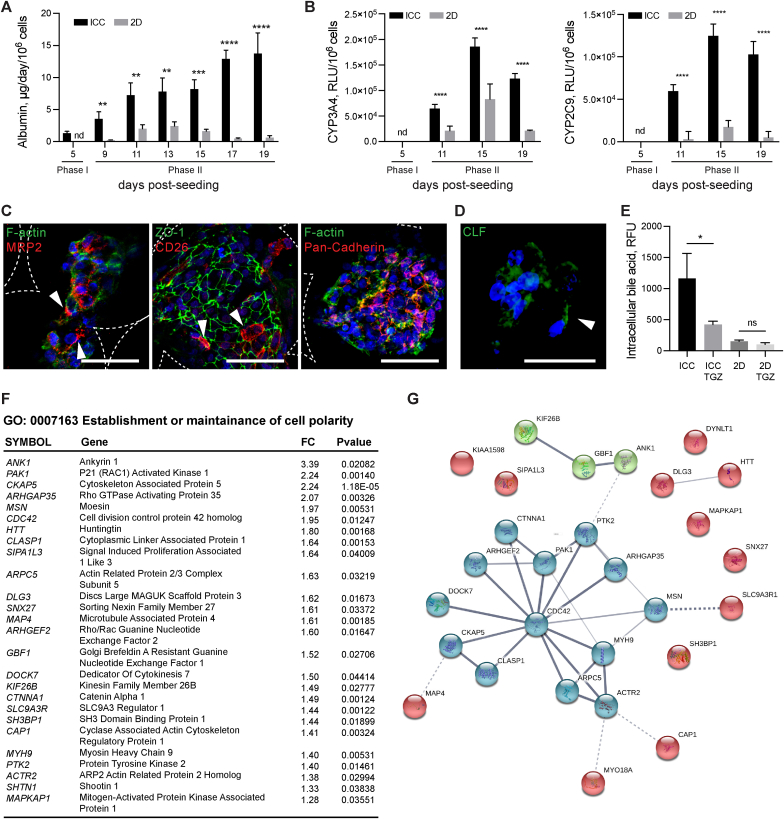
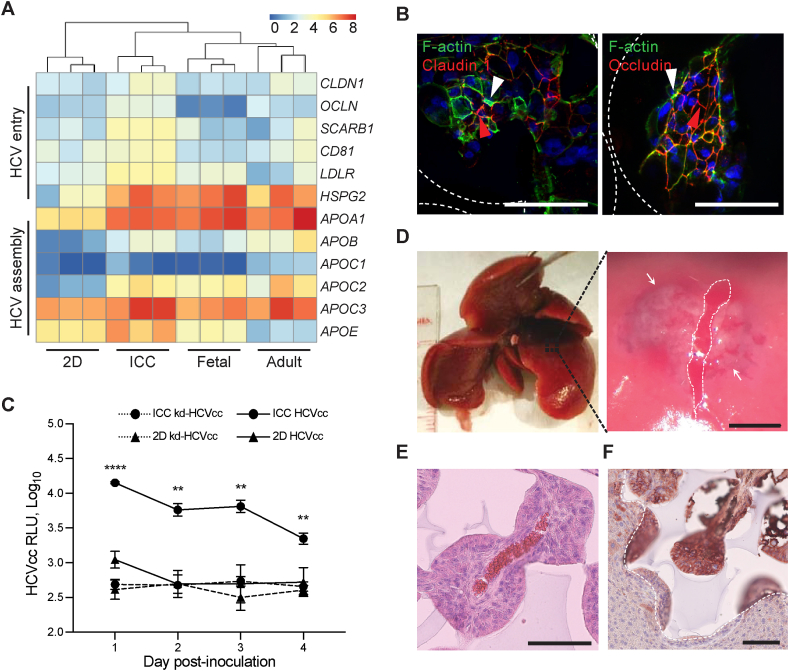
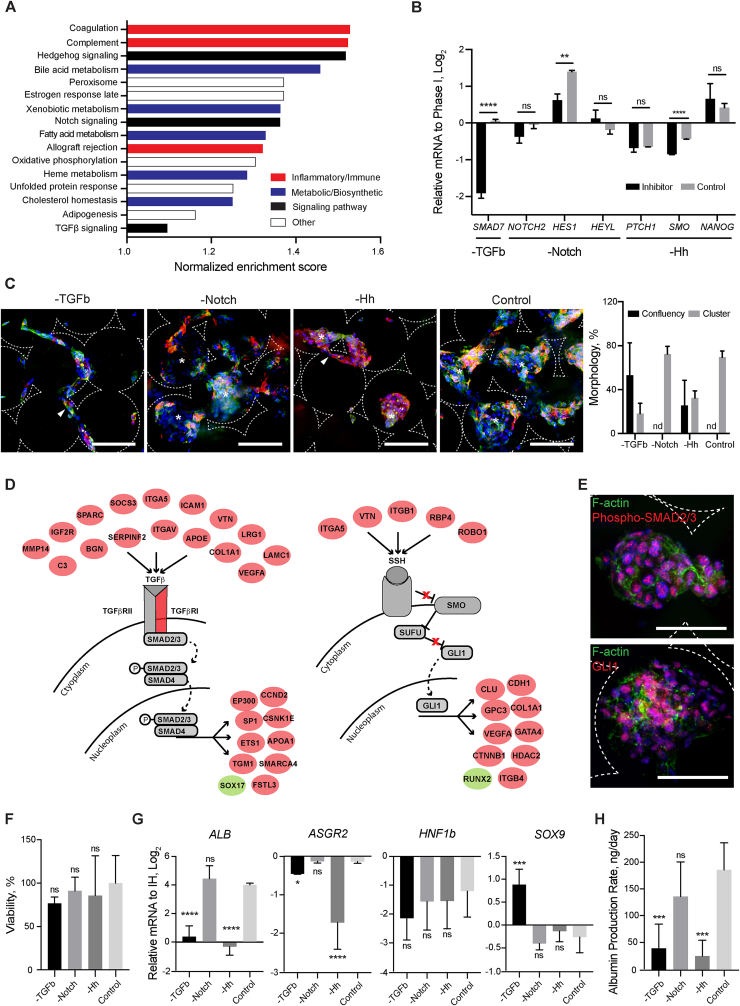
Generation of human organoids from induced pluripotent stem cells (iPSCs) offers exciting possibilities for developmental biology, disease modelling and cell therapy. Significant advances towards those goals have been hampered by dependence on animal derived matrices (e.g. Matrigel), immortalized cell lines and resultant structures that are difficult to control or scale. To address these challenges, we aimed to develop a fully defined liver organoid platform using inverted colloid crystal (ICC) whose 3-dimensional mechanical properties could be engineered to recapitulate the extracellular niche sensed by hepatic progenitors during human development. iPSC derived hepatic progenitors (IH) formed organoids most optimally in ICC scaffolds constructed with 140 μm diameter pores coated with type I collagen in a two-step process mimicking liver bud formation. The resultant organoids were closer to adult tissue, compared to 2D and 3D controls, with respect to morphology, gene expression, protein secretion, drug metabolism and viral infection and could integrate, vascularise and function following implantation into livers of immune-deficient mice. Preliminary interrogation of the underpinning mechanisms highlighted the importance of TGFβ and hedgehog signalling pathways. The combination of functional relevance with tuneable mechanical properties leads us to propose this bioengineered platform to be ideally suited for a range of future mechanistic and clinical organoid related applications.
Keywords: Bioengineering; Biomimetic materials; Liver stem cells; Organogenesis.
Copyright © 2018 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Takebe T. Vascularized and functional human liver from an iPSC-derived organ bud transplant. Nature. 2013;499(7459):481–484. - PubMed
-
- Sato T. Single Lgr5 stem cells build crypt–villus structures in vitro without a mesenchymal niche. Nature. 2009;459(7244):262–265. - PubMed
-
- Takasato M. Kidney organoids from human iPS cells contain multiple lineages and model human nephrogenesis. Nature. 2015;526(7574):564–568. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
