

Determining the molecular basis of voltage sensitivity in membrane proteins
- PMID: 30150239
- PMCID: PMC6168238
- DOI: 10.1085/jgp.201812086
Determining the molecular basis of voltage sensitivity in membrane proteins
Abstract
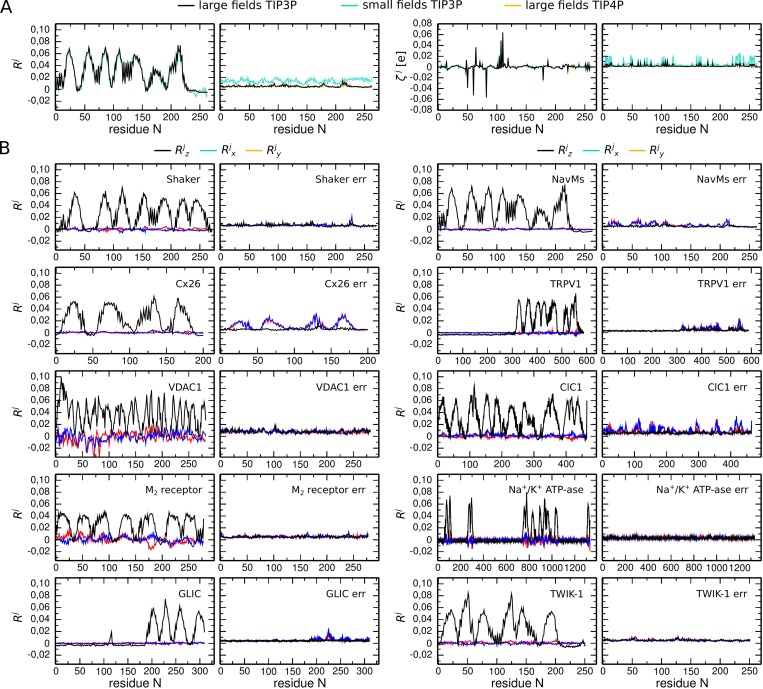
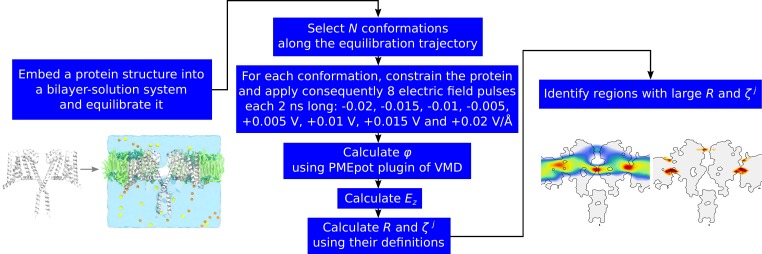
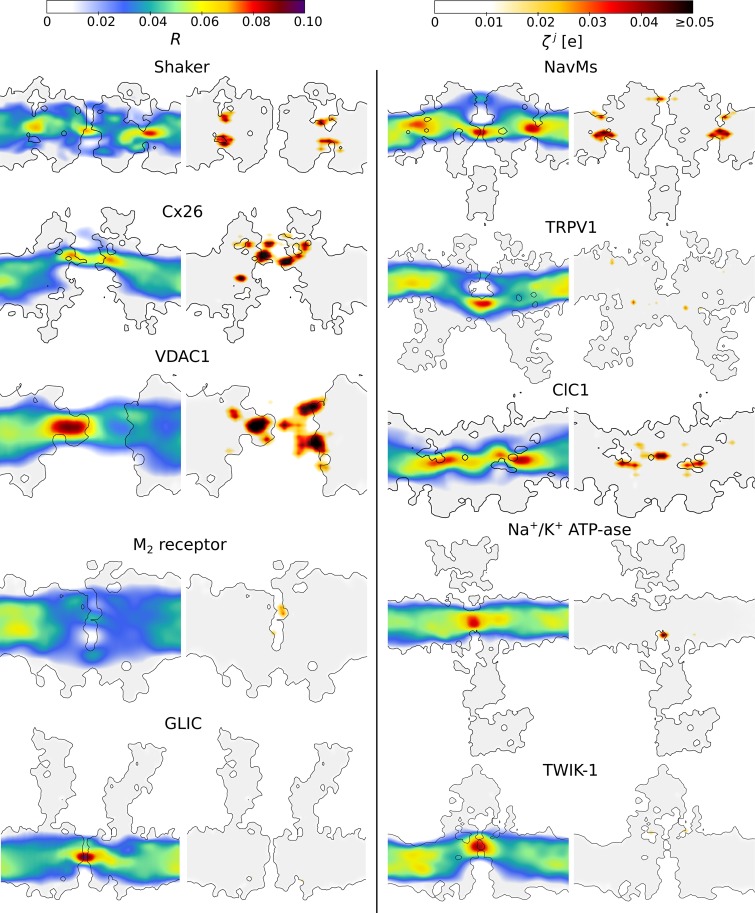
Voltage-sensitive membrane proteins are united by their ability to transform changes in membrane potential into mechanical work. They are responsible for a spectrum of physiological processes in living organisms, including electrical signaling and cell-cycle progression. Although the mechanism of voltage-sensing has been well characterized for some membrane proteins, including voltage-gated ion channels, even the location of the voltage-sensing elements remains unknown for others. Moreover, the detection of these elements by using experimental techniques is challenging because of the diversity of membrane proteins. Here, we provide a computational approach to predict voltage-sensing elements in any membrane protein, independent of its structure or function. It relies on an estimation of the propensity of a protein to respond to changes in membrane potential. We first show that this property correlates well with voltage sensitivity by applying our approach to a set of voltage-sensitive and voltage-insensitive membrane proteins. We further show that it correctly identifies authentic voltage-sensitive residues in the voltage-sensor domain of voltage-gated ion channels. Finally, we investigate six membrane proteins for which the voltage-sensing elements have not yet been characterized and identify residues and ions that might be involved in the response to voltage. The suggested approach is fast and simple and enables a characterization of voltage sensitivity that goes beyond mere identification of charges. We anticipate that its application before mutagenesis experiments will significantly reduce the number of potential voltage-sensitive elements to be tested.
© 2018 Kasimova et al.
Figures
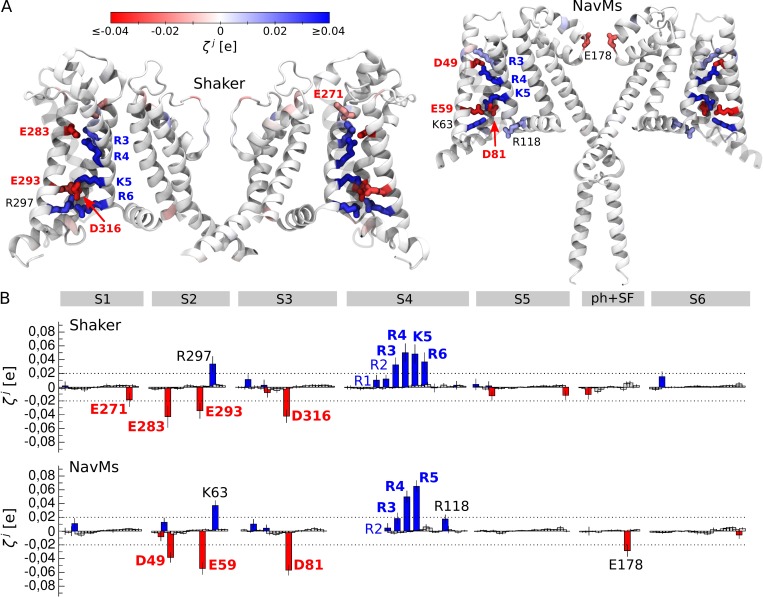
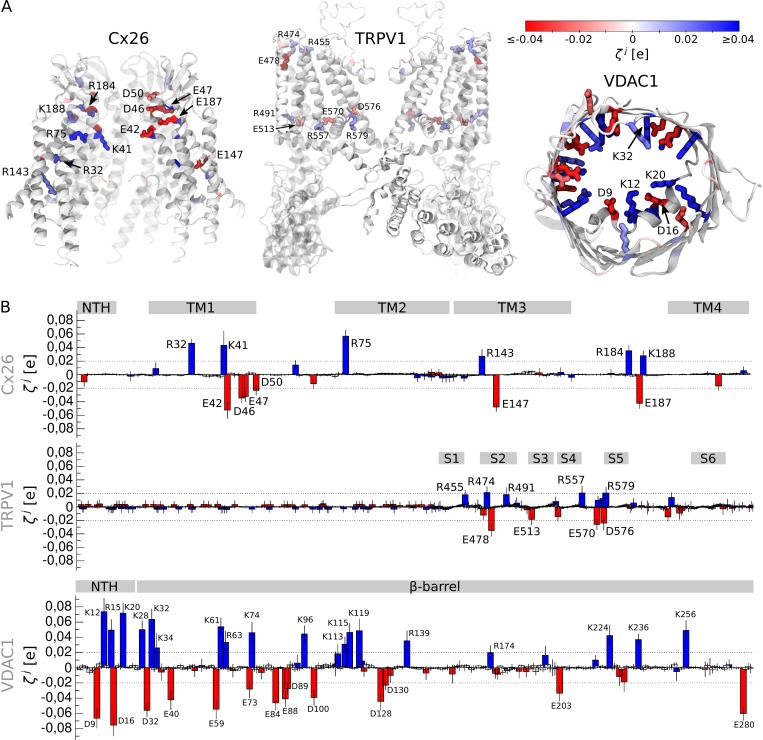
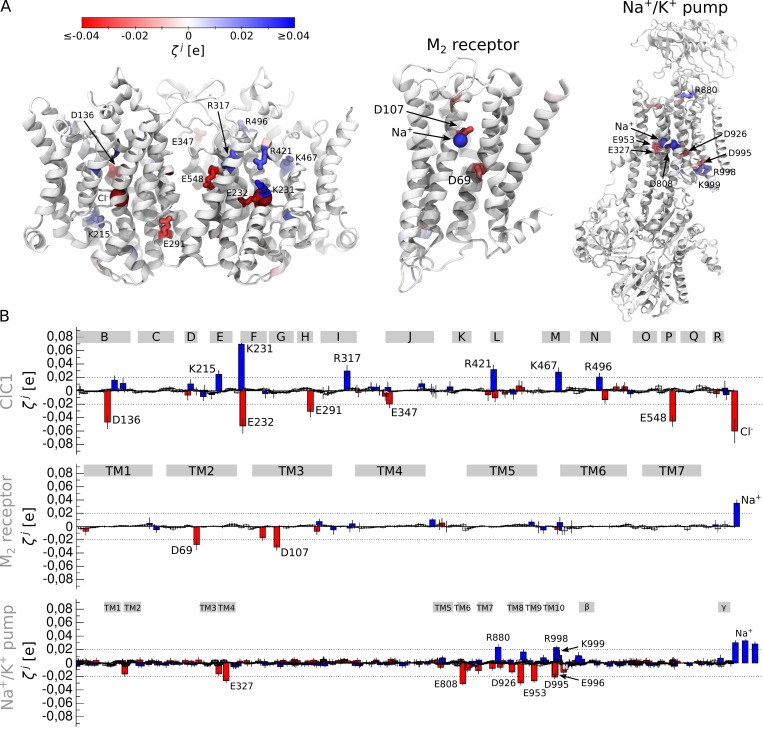
Comment in
-
Predicting voltage sensing.J Gen Physiol. 2018 Oct 1;150(10):1349. doi: 10.1085/jgp.201812233. Epub 2018 Sep 10. J Gen Physiol. 2018. PMID: 30201729 Free PMC article.
References
-
- Abraham M.J., Murtola T., Schulz R., Páll S., Smith J.C., Hess B., and Lindahl E.. 2015. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX. 1–2:19–25. 10.1016/j.softx.2015.06.001 - DOI
-
- Anzai N., Ichida K., Jutabha P., Kimura T., Babu E., Jin C.J., Srivastava S., Kitamura K., Hisatome I., Endou H., and Sakurai H.. 2008. Plasma urate level is directly regulated by a voltage-driven urate efflux transporter URATv1 (SLC2A9) in humans. J. Biol. Chem. 283:26834–26838. 10.1074/jbc.C800156200 - DOI - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
