

Epidermal Growth Factor (EGF) Autocrine Activation of Human Platelets Promotes EGF Receptor-Dependent Oral Squamous Cell Carcinoma Invasion, Migration, and Epithelial Mesenchymal Transition
- PMID: 30150285
- PMCID: PMC6143412
- DOI: 10.4049/jimmunol.1800124
Epidermal Growth Factor (EGF) Autocrine Activation of Human Platelets Promotes EGF Receptor-Dependent Oral Squamous Cell Carcinoma Invasion, Migration, and Epithelial Mesenchymal Transition
Abstract
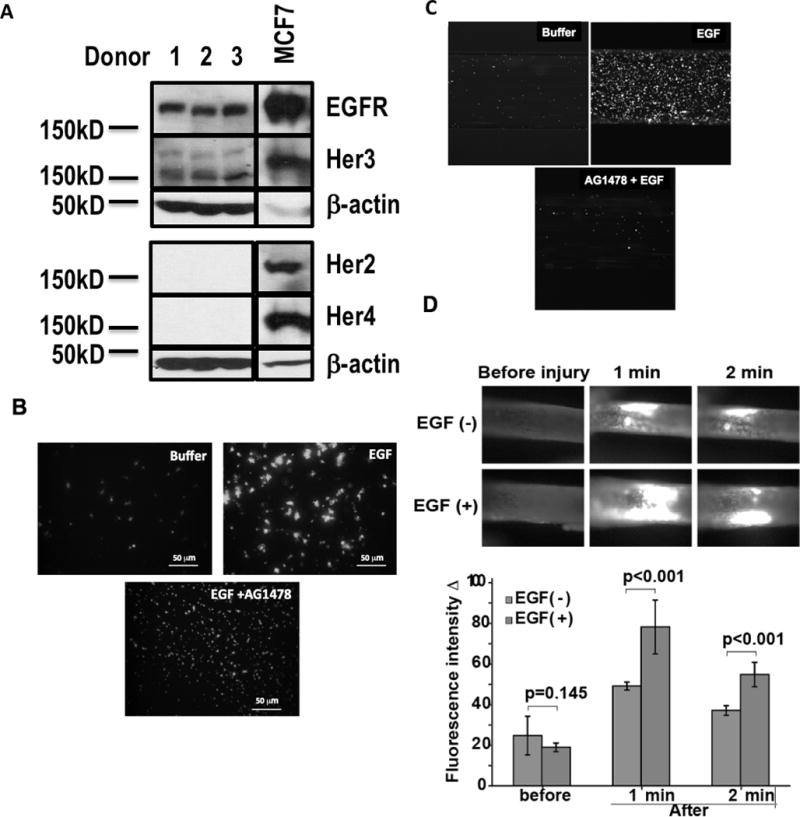
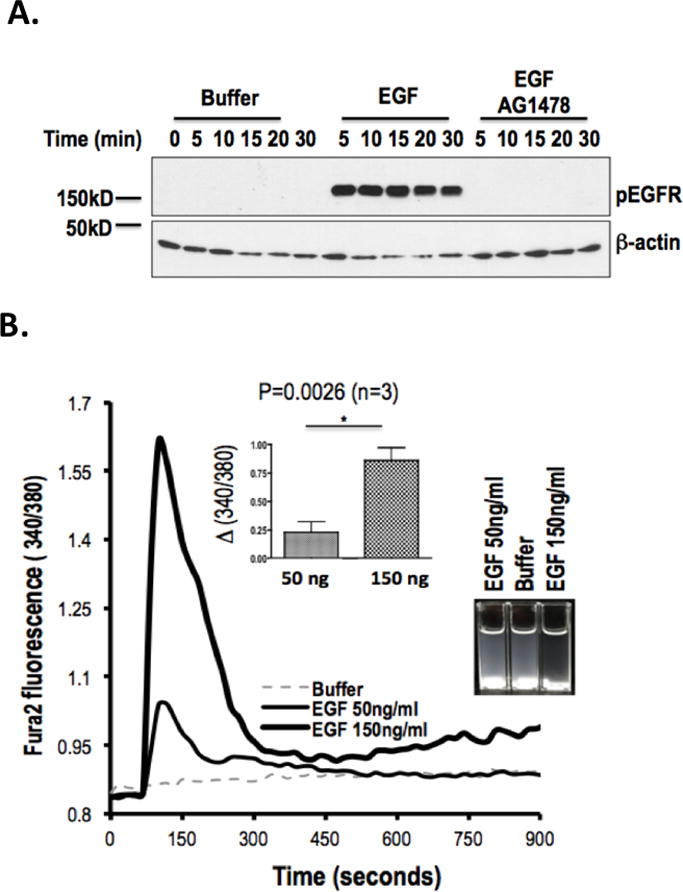
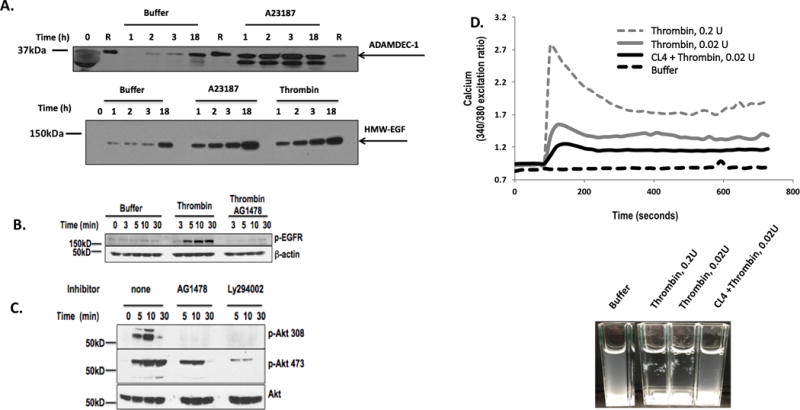
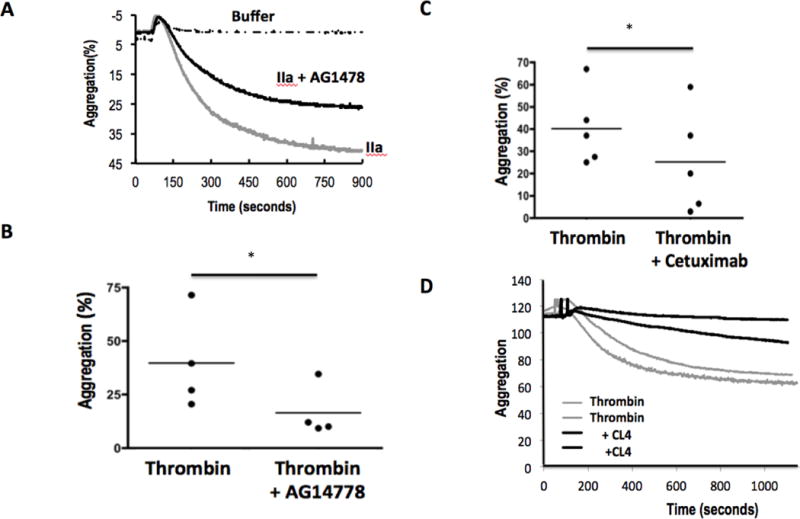
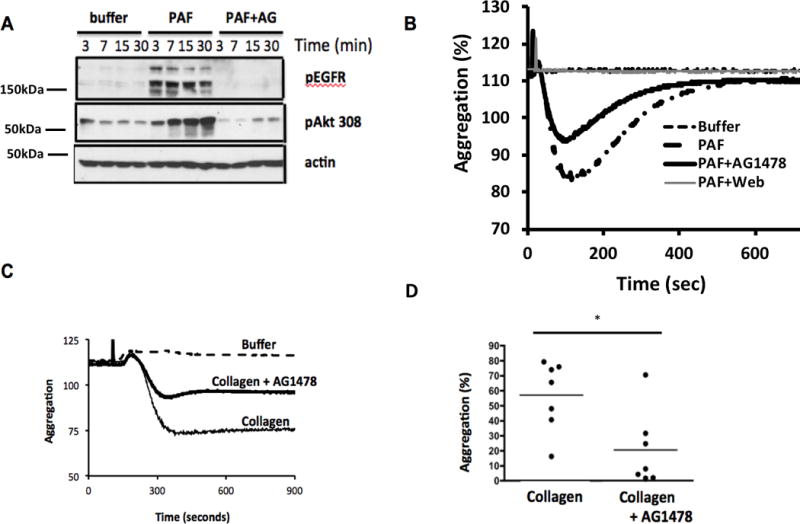
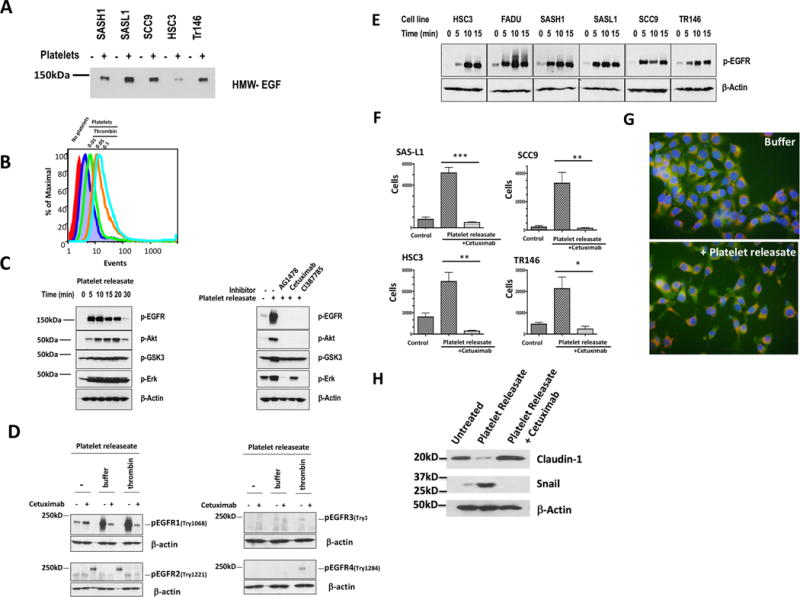
Activated platelets release functional, high m.w. epidermal growth factor (HMW-EGF). In this study, we show platelets also express epidermal growth factor (EGF) receptor (EGFR) protein, but not ErbB2 or ErbB4 coreceptors, and so might respond to HMW-EGF. We found HMW-EGF stimulated platelet EGFR autophosphorylation, PI3 kinase-dependent AKT phosphorylation, and a Ca2+ transient that were blocked by EGFR tyrosine kinase inhibition. Strong (thrombin) and weak (ADP, platelet-activating factor) G protein-coupled receptor agonists and non-G protein-coupled receptor collagen recruited EGFR tyrosine kinase activity that contributed to platelet activation because EGFR kinase inhibition reduced signal transduction and aggregation induced by each agonist. EGF stimulated ex vivo adhesion of platelets to collagen-coated microfluidic channels, whereas systemic EGF injection increased initial platelet deposition in FeCl3-damaged murine carotid arteries. EGFR signaling contributes to oral squamous cell carcinoma (OSCC) tumorigenesis, but the source of its ligand is not established. We find individual platelets were intercalated within OSCC tumors. A portion of these platelets expressed stimulation-dependent Bcl-3 and IL-1β and so had been activated. Stimulated platelets bound OSCC cells, and material released from stimulated platelets induced OSCC epithelial-mesenchymal transition and stimulated their migration and invasion through Matrigel barriers. Anti-EGF Ab or EGFR inhibitors abolished platelet-induced tumor cell phenotype transition, migration, and invasion; so the only factor released from activated platelets necessary for OSCC metastatic activity was HMW-EGF. These results establish HMW-EGF in platelet function and elucidate a previously unsuspected connection between activated platelets and tumorigenesis through rapid, and prolonged, autocrine-stimulated release of HMW-EGF by tumor-associated platelets.
Copyright © 2018 by The American Association of Immunologists, Inc.
Conflict of interest statement
Disclosures
The authors have no financial conflicts of interest.
Figures

References
-
- van Holten TC, Bleijerveld OB, Wijten P, de Groot PG, Heck AJ, Barendrecht AD, Merkx TH, Scholten A, Roest M. Quantitative proteomics analysis reveals similar release profiles following specific PAR-1 or PAR-4 stimulation of platelets. Cardiovasc Res. 2014;103:140–146. - PubMed
-
- Wijten P, van Holten T, Woo LL, Bleijerveld OB, Roest M, Heck AJ, Scholten A. High precision platelet releasate definition by quantitative reversed protein profiling–brief report. Arterioscler Thromb Vasc Biol. 2013;33:1635–1638. - PubMed
-
- Semple JW, Italiano JE, Jr, Freedman J. Platelets and the immune continuum. Nat Rev Immunol. 2011;11:264–274. - PubMed
-
- Su CY, Kuo YP, Nieh HL, Tseng YH, Burnouf T. Quantitative assessment of the kinetics of growth factors release from platelet gel. Transfusion (Paris) 2008;48:2414–2420. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
