

Conformational changes in Arp2/3 complex induced by ATP, WASp-VCA, and actin filaments
- PMID: 30150414
- PMCID: PMC6140485
- DOI: 10.1073/pnas.1717594115
Conformational changes in Arp2/3 complex induced by ATP, WASp-VCA, and actin filaments
Abstract
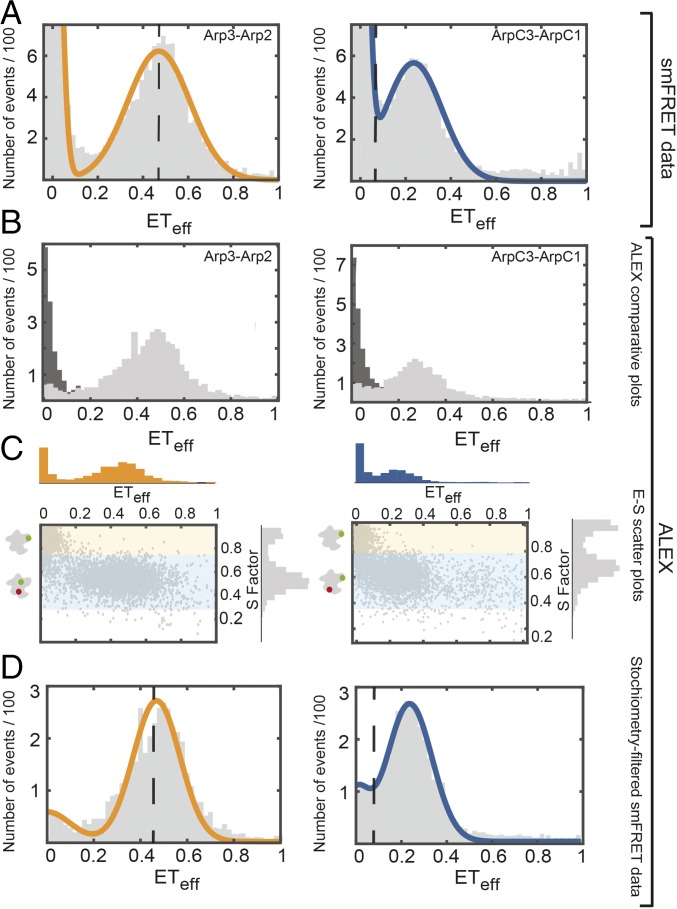
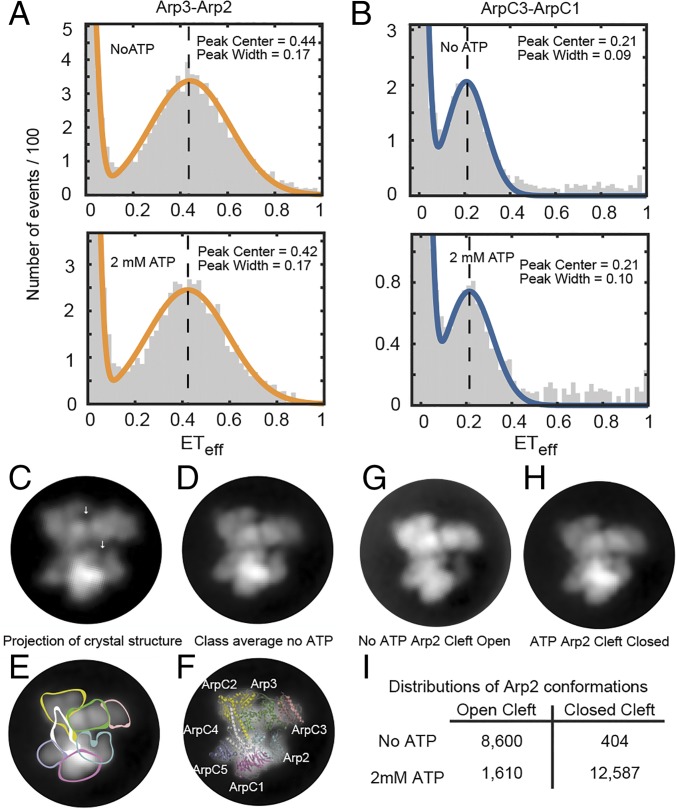
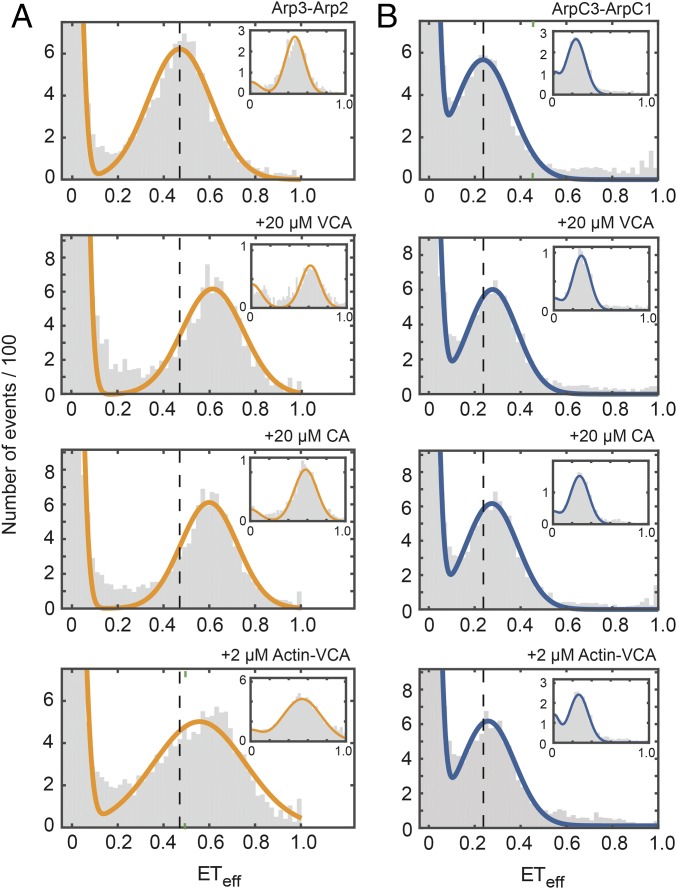
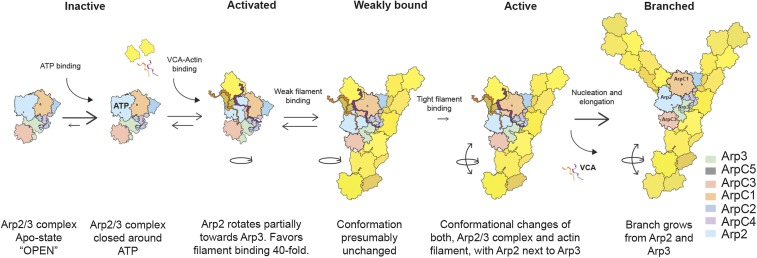
We used fluorescence spectroscopy and EM to determine how binding of ATP, nucleation-promoting factors, actin monomers, and actin filaments changes the conformation of Arp2/3 complex during the process that nucleates an actin filament branch. We mutated subunits of Schizosaccharomyces pombe Arp2/3 complex for labeling with fluorescent dyes at either the C termini of Arp2 and Arp3 or ArpC1 and ArpC3. We measured Förster resonance energy transfer (FRET) efficiency (ETeff) between the dyes in the presence of the various ligands. We also computed class averages from electron micrographs of negatively stained specimens. ATP binding made small conformational changes of the nucleotide-binding cleft of the Arp2 subunit. WASp-VCA, WASp-CA, and WASp-actin-VCA changed the ETeff between the dyes on the Arp2 and Arp3 subunits much more than between dyes on ArpC1 and ArpC3. Ensemble FRET detected an additional structural change that brought ArpC1 and ArpC3 closer together when Arp2/3 complex bound actin filaments. VCA binding to Arp2/3 complex causes a conformational change that favors binding to the side of an actin filament, which allows further changes required to nucleate a daughter filament.
Keywords: Arp2/3 complex; WASp; actin; electron microscopy; fluorescence.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Pollard TD, Borisy GG. Cellular motility driven by assembly and disassembly of actin filaments. Cell. 2003;112:453–465. - PubMed
-
- Goley ED, Welch MD. The ARP2/3 complex: An actin nucleator comes of age. Nat Rev Mol Cell Biol. 2006;7:713–726. - PubMed
-
- Pollard TD, Blanchoin L, Mullins RD. Molecular mechanisms controlling actin filament dynamics in nonmuscle cells. Annu Rev Biophys Biomol Struct. 2000;29:545–576. - PubMed
-
- Chhabra ES, Higgs HN. The many faces of actin: Matching assembly factors with cellular structures. Nat Cell Biol. 2007;9:1110–1121. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
