

Insulin-Like Growth Factor-I as an Effector Element of the Cytokine IL-4 in the Development of a Leishmania major Infection
- PMID: 30150896
- PMCID: PMC6087592
- DOI: 10.1155/2018/9787128
Insulin-Like Growth Factor-I as an Effector Element of the Cytokine IL-4 in the Development of a Leishmania major Infection
Abstract
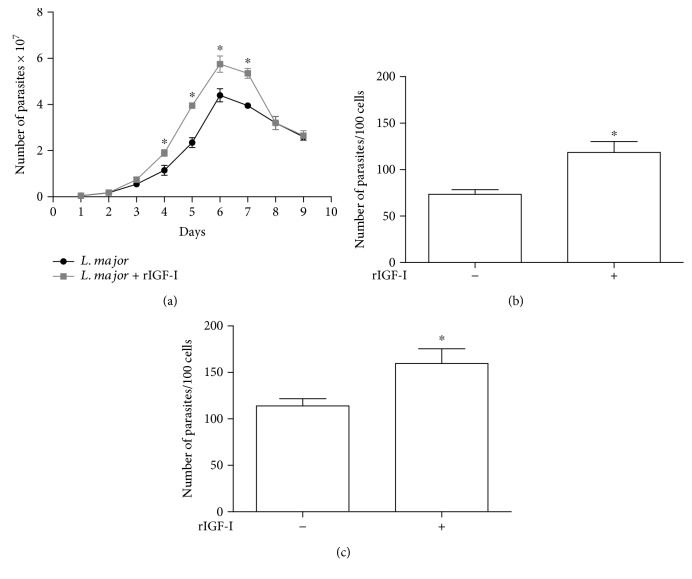
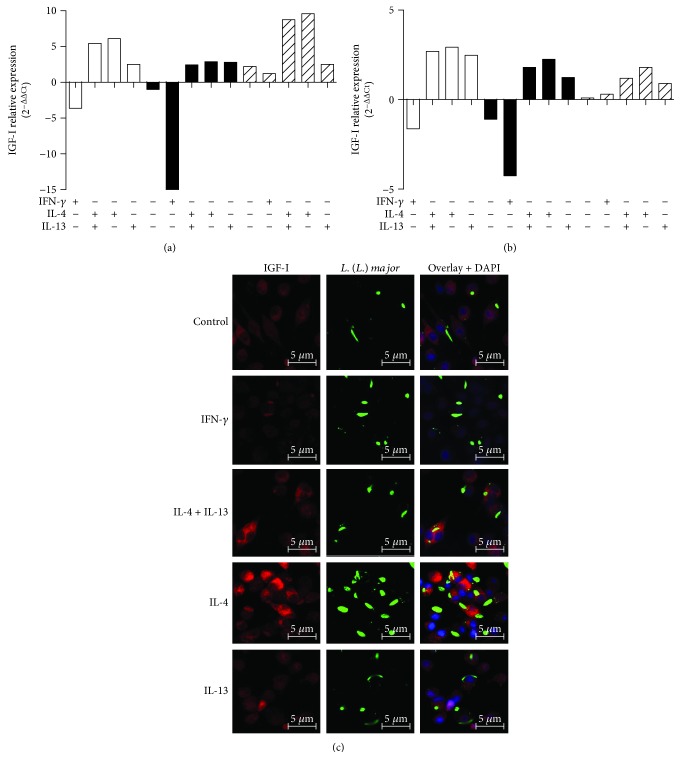
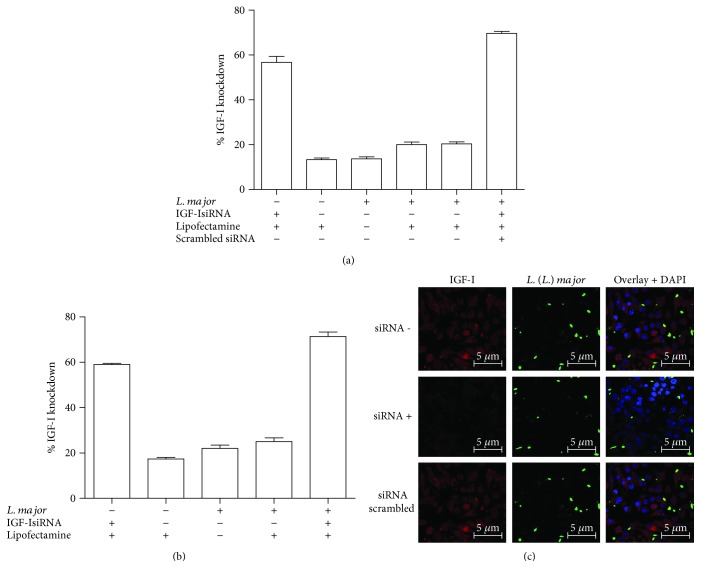
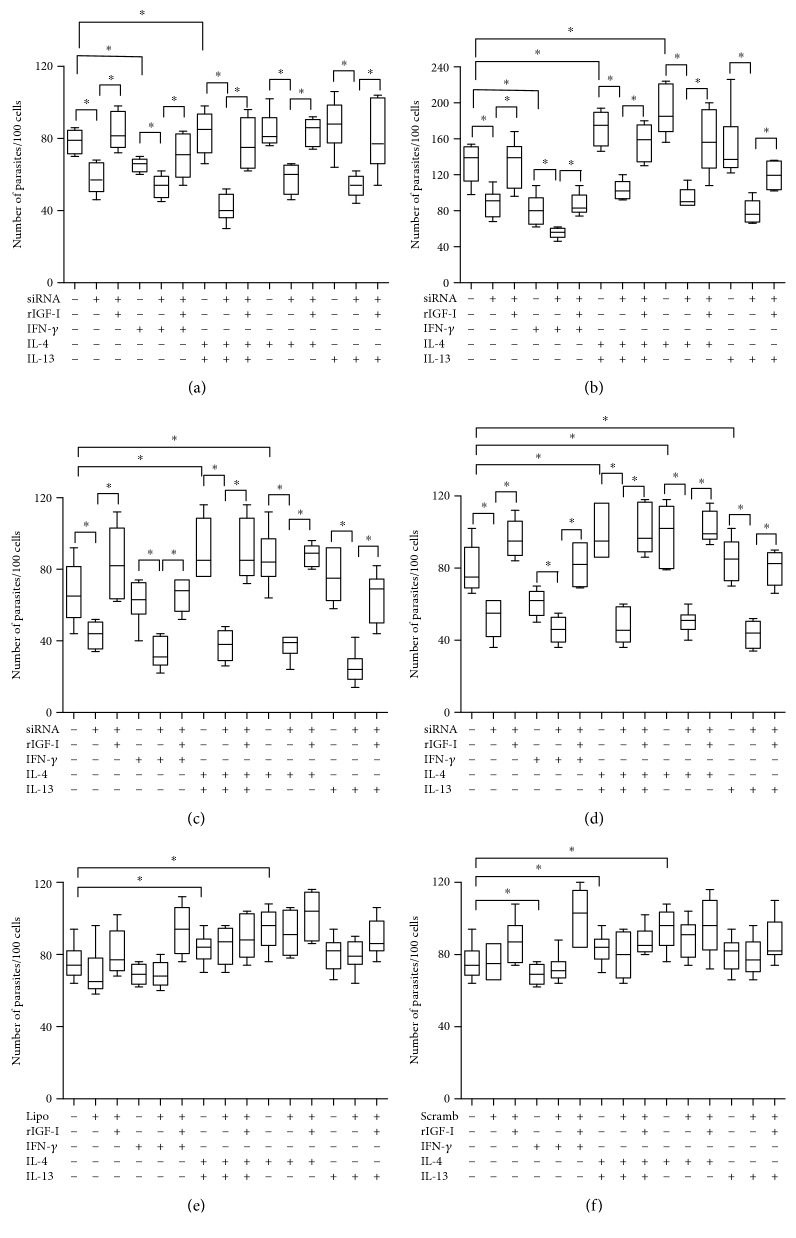
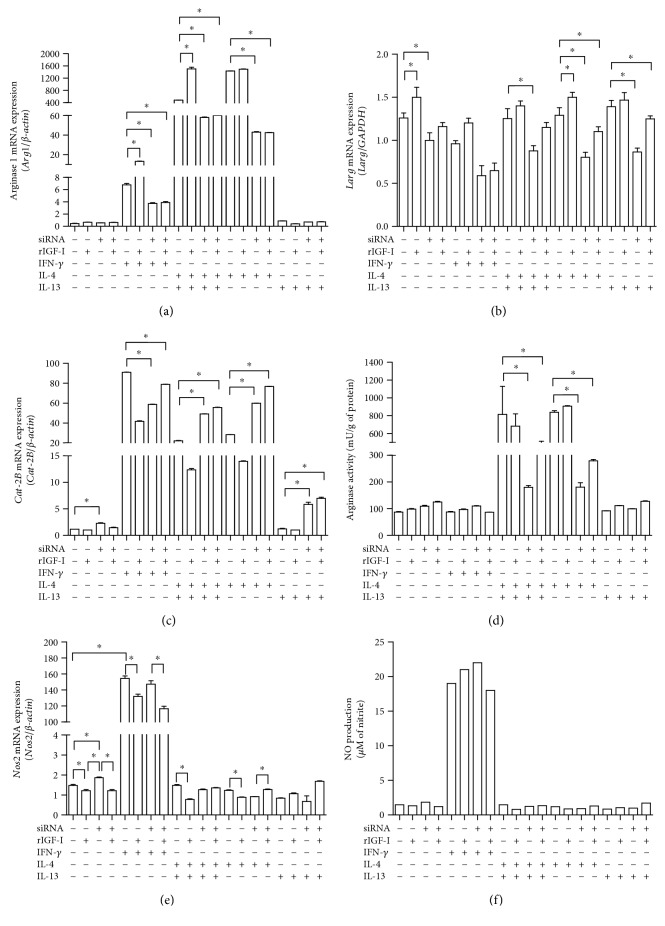
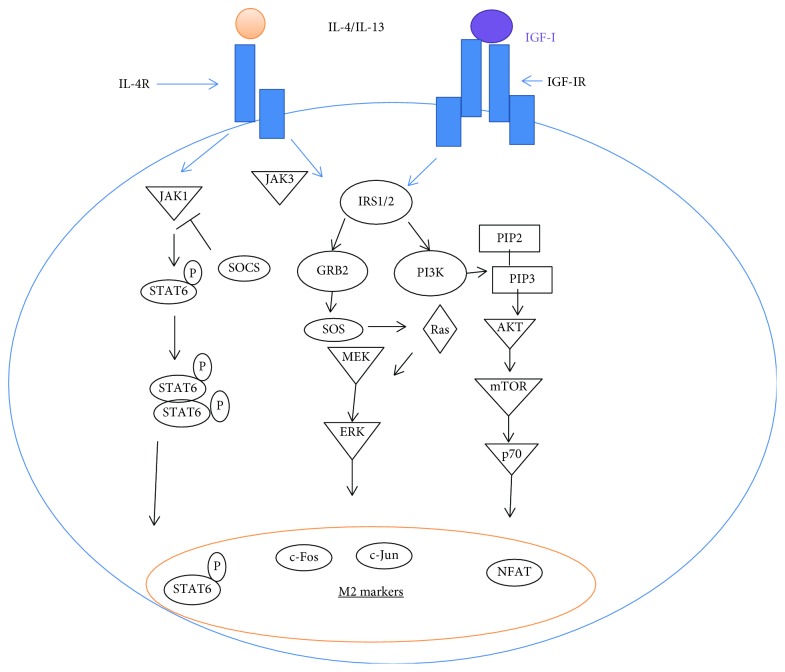
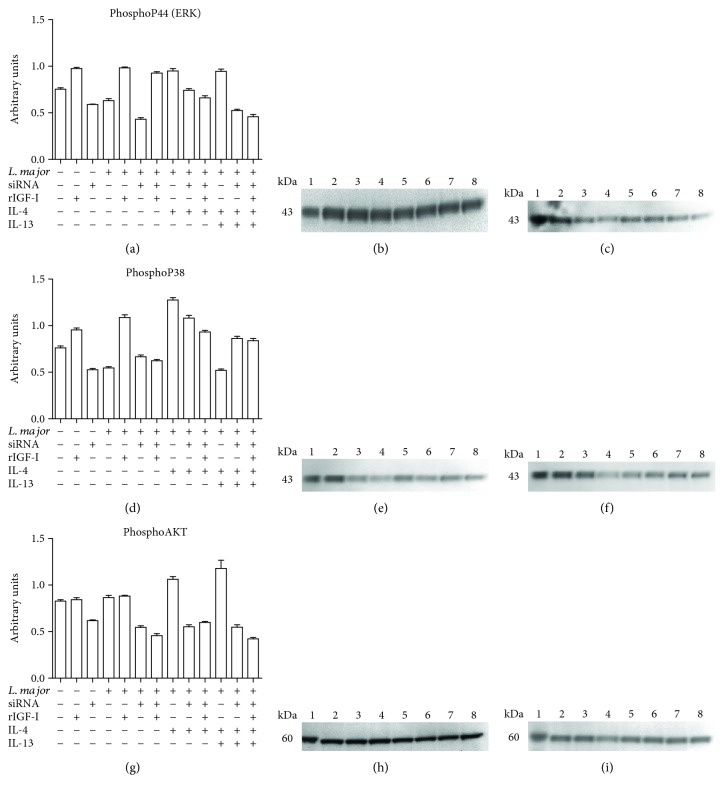
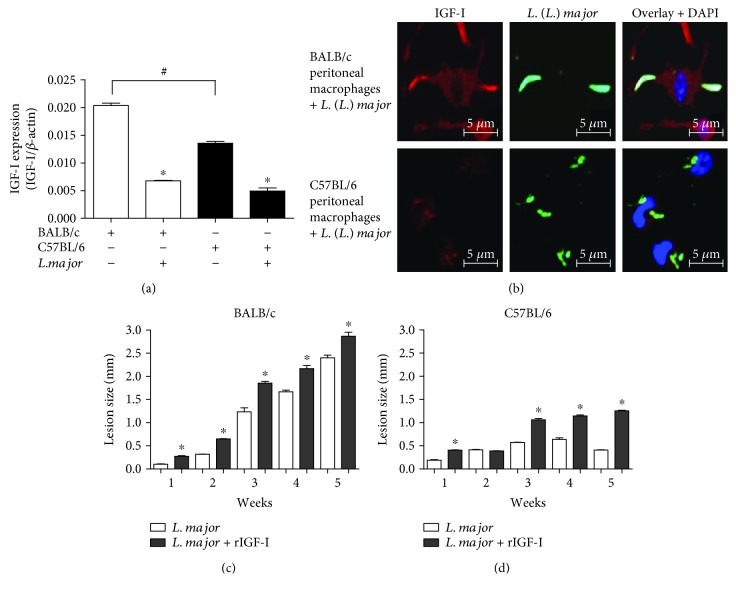
Certain cytokines modulate the expression of insulin-like growth factor- (IGF-) I. Since IL-4 and IGF-I promote growth of the protozoan Leishmania major, we here addressed their interaction in downregulating the expression of Igf-I mRNA using small interfering RNA (siRNA) in Leishmania major-infected macrophages. Parasitism was decreased in the siRNA-treated cells compared with the nontreated cells, reversed by the addition of recombinant IGF-I (rIGF-I). In IL-4-stimulated macrophages, parasitism and the Igf-I mRNA amount were increased, and the effects were nullified upon siRNA transfection. IGF-I downregulation inhibited both parasite and macrophage arginase activation even in IL-4-stimulated cells. Searching for intracellular signaling components shared by IL-4 and IGF-I, upon siRNA transfection, phosphorylated p44, p38, and Akt proteins were decreased, affecting the phosphatidylinositol-3-kinase (PI3K)/Akt pathway. In L. major-infected C57BL6-resistant mice, the preincubation of the parasite with rIGF-I changed the infection profile to be similar to that of susceptible mice. We conclude that IGF-I constitutes an effector element of IL-4 involving the PI3K/Akt pathway during L. major infection.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
