

The emerging roles of ribosomal histidyl hydroxylases in cell biology, physiology and disease
- PMID: 30151692
- PMCID: PMC6182338
- DOI: 10.1007/s00018-018-2903-z
The emerging roles of ribosomal histidyl hydroxylases in cell biology, physiology and disease
Abstract
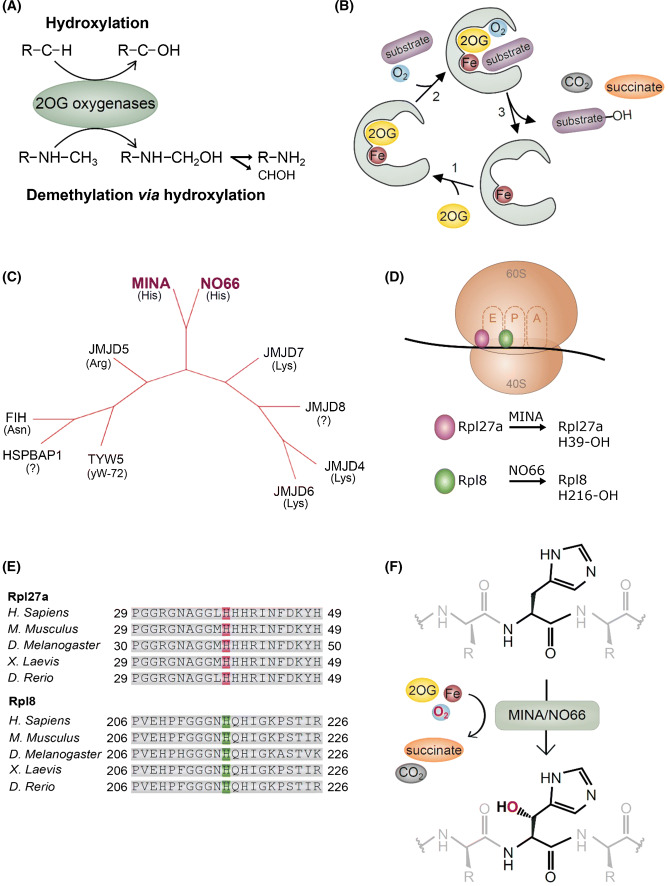
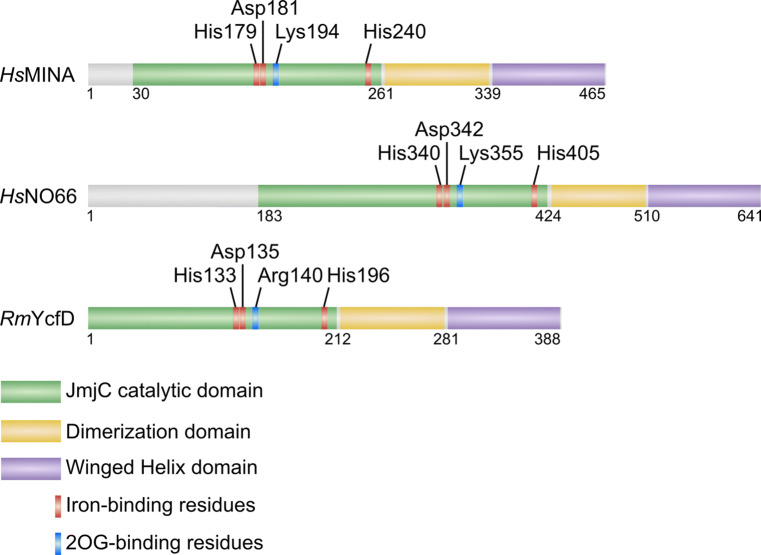
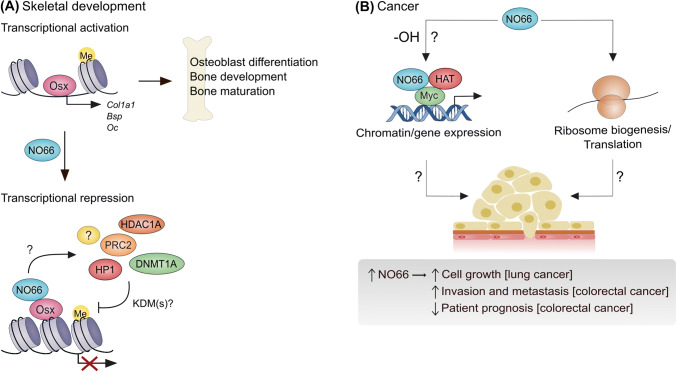
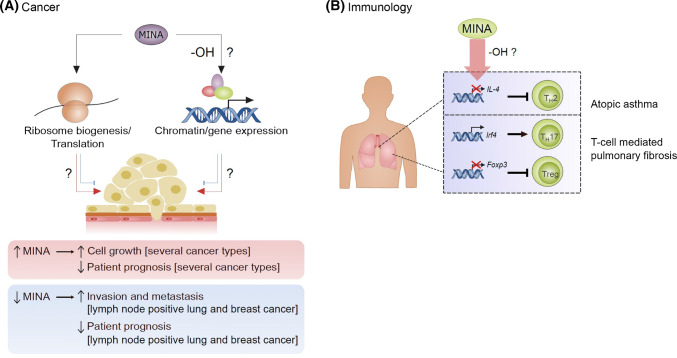
Hydroxylation is a novel protein modification catalyzed by a family of oxygenases that depend on fundamental nutrients and metabolites for activity. Protein hydroxylases have been implicated in a variety of key cellular processes that play important roles in both normal homeostasis and pathogenesis. Here, in this review, we summarize the current literature on a highly conserved sub-family of oxygenases that catalyze protein histidyl hydroxylation. We discuss the evidence supporting the biochemical assignment of these emerging enzymes as ribosomal protein hydroxylases, and provide an overview of their role in immunology, bone development, and cancer.
Keywords: Bone development; Cancer; Histidine; Hydroxylation; Immunology; Post-translational modification.
Conflict of interest statement
The authors report no conflicts of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
