

Identification and expression analysis of putative chemoreception genes from Cyrtorhinus lividipennis (Hemiptera: Miridae) antennal transcriptome
- PMID: 30154418
- PMCID: PMC6113244
- DOI: 10.1038/s41598-018-31294-9
Identification and expression analysis of putative chemoreception genes from Cyrtorhinus lividipennis (Hemiptera: Miridae) antennal transcriptome
Abstract
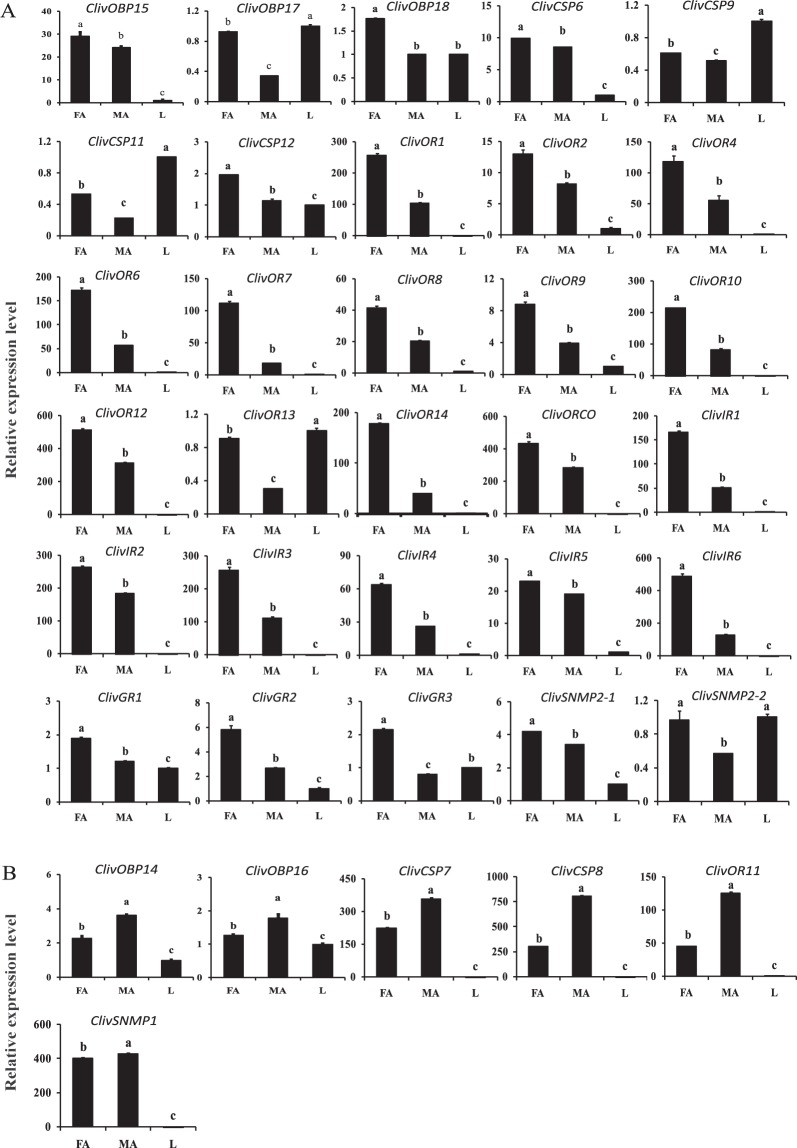
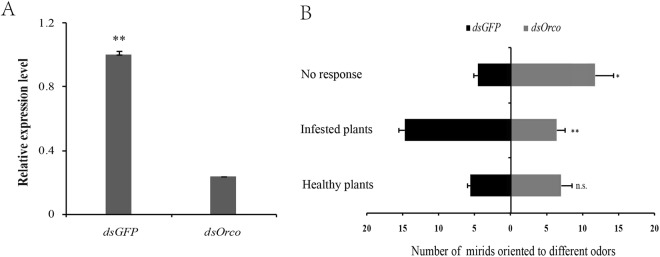
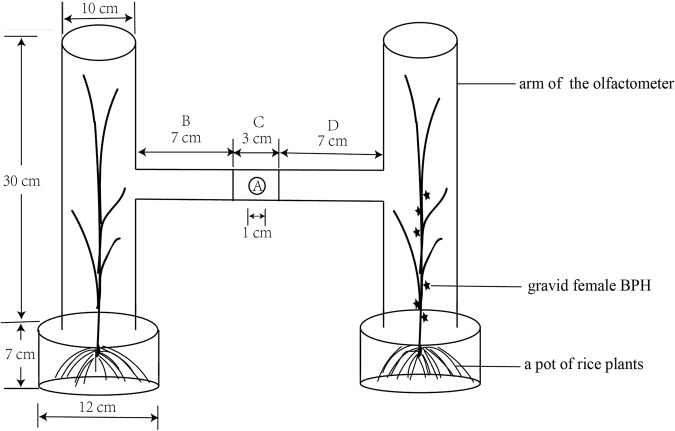
Cyrtorhinus lividipennis Reuter (Hemiptera: Miridae) is an important egg predator of planthoppers which are destructive rice pests. The chemosensory genes in the mirid antennae play important roles in mating and prey-seeking behaviors. To gain a better understanding of the olfaction of C. lividipennis, we sequenced the antennal transcriptomes of the predator to identify the key olfaction genes. We identified 18 odorant binding proteins (OBPs), 12 chemosensory proteins (CSPs), 1 Niemann-Pick C2 protein (NPC2), 15 odorant receptors (ORs), 6 ionotropic receptors (IRs), 3 gustatory receptors (GRs) and 3 sensory neuron membrane proteins (SNMPs). Quantitative real-time PCR results showed that the relative transcript levels of three ClivORs (ClivOR6, 7 and 14) in the female antennae were 3 to 6 folds higher than that in the male antennae, indicating these genes were more related to oviposition site selection. The relative transcript levels of ClivCSP8 and ClivOR11 were 2.6 and 2.7 times higher in the male antennae than that of the female, respectively, indicating that these genes might be involved in mate searching. Moreover, the responses of dsorco treated predators to volatiles emitted from infested rice were significantly reduced, indicating these volatiles might serve as crucial cues in the host searching of C. lividipennis.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Identification of Candidate Odorant-Binding Protein and Chemosensory Protein Genes in Cyrtorhinus lividipennis (Hemiptera: Miridae), a Key Predator of the Rice Planthoppers in Asia.Environ Entomol. 2017 Jun 1;46(3):654-662. doi: 10.1093/ee/nvx075. Environ Entomol. 2017. PMID: 28407047
-
Antennal and Abdominal Transcriptomes Reveal Chemosensory Genes in the Asian Citrus Psyllid, Diaphorina citri.PLoS One. 2016 Jul 21;11(7):e0159372. doi: 10.1371/journal.pone.0159372. eCollection 2016. PLoS One. 2016. PMID: 27441376 Free PMC article.
-
Identification and characterization of chemosensory genes in the antennal transcriptome of Spodoptera exigua.Comp Biochem Physiol Part D Genomics Proteomics. 2018 Sep;27:54-65. doi: 10.1016/j.cbd.2018.05.001. Epub 2018 May 7. Comp Biochem Physiol Part D Genomics Proteomics. 2018. PMID: 29787920
-
Identification of candidate olfactory genes in cicada Subpsaltria yangi by antennal transcriptome analysis.Comp Biochem Physiol Part D Genomics Proteomics. 2018 Dec;28:122-133. doi: 10.1016/j.cbd.2018.08.001. Epub 2018 Sep 1. Comp Biochem Physiol Part D Genomics Proteomics. 2018. PMID: 30195211
-
Identification and characterization of the distinct expression profiles of candidate chemosensory membrane proteins in the antennal transcriptome of Adelphocoris lineolatus (Goeze).Insect Mol Biol. 2017 Feb;26(1):74-91. doi: 10.1111/imb.12272. Epub 2016 Nov 26. Insect Mol Biol. 2017. PMID: 27888648
Cited by
-
Comparative transcriptomic analysis of salivary glands between the zoophytophagous Cyrtorhinus lividipennis and the phytozoophagous Apolygus lucorum.BMC Genomics. 2024 Jan 11;25(1):53. doi: 10.1186/s12864-023-09956-4. BMC Genomics. 2024. PMID: 38212677 Free PMC article.
-
Antennal Transcriptome Analysis and Identification of Candidate Chemosensory Genes of the Harlequin Ladybird Beetle, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae).Insects. 2021 Mar 2;12(3):209. doi: 10.3390/insects12030209. Insects. 2021. PMID: 33801288 Free PMC article.
-
Identification and characterization of soluble binding proteins associated with host foraging in the parasitoid wasp Diachasmimorpha longicaudata.PLoS One. 2021 Jun 17;16(6):e0252765. doi: 10.1371/journal.pone.0252765. eCollection 2021. PLoS One. 2021. PMID: 34138896 Free PMC article.
-
The pivotal roles of gut microbiota in insect plant interactions for sustainable pest management.NPJ Biofilms Microbiomes. 2023 Sep 21;9(1):66. doi: 10.1038/s41522-023-00435-y. NPJ Biofilms Microbiomes. 2023. PMID: 37735530 Free PMC article. Review.
-
Identification and odor exposure regulation of odorant-binding proteins in Picromerus lewisi.Front Physiol. 2024 Dec 4;15:1503440. doi: 10.3389/fphys.2024.1503440. eCollection 2024. Front Physiol. 2024. PMID: 39697614 Free PMC article.
References
-
- Ponzio C, et al. Volatile-mediated foraging behaviour of three parasitoid species under conditions of dual insect herbivore attack. Anim. Behav. 2016;111:197–206. doi: 10.1016/j.anbehav.2015.10.024. - DOI
-
- Heong KL, Bleih S, Lazaro AA. Predation of of Cyrtorhinus lividipennis Reuter on eggs of the green leafhopper and brown planthopper in rice. Res. Popul. Ecol. 1990;32:255–262. doi: 10.1007/BF02512561. - DOI
-
- Qiao F, et al. Reciprocal intraguild predation between two mirid predators, Cyrtorhinus lividipennis and Tytthus chinensis (Hemiptera: Miridae) Biocontrol Sci. Techn. 2016;26:1267–1284. doi: 10.1080/09583157.2016.1194370. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
