

Psychophysical reverse correlation reflects both sensory and decision-making processes
- PMID: 30154467
- PMCID: PMC6113286
- DOI: 10.1038/s41467-018-05797-y
Psychophysical reverse correlation reflects both sensory and decision-making processes
Abstract
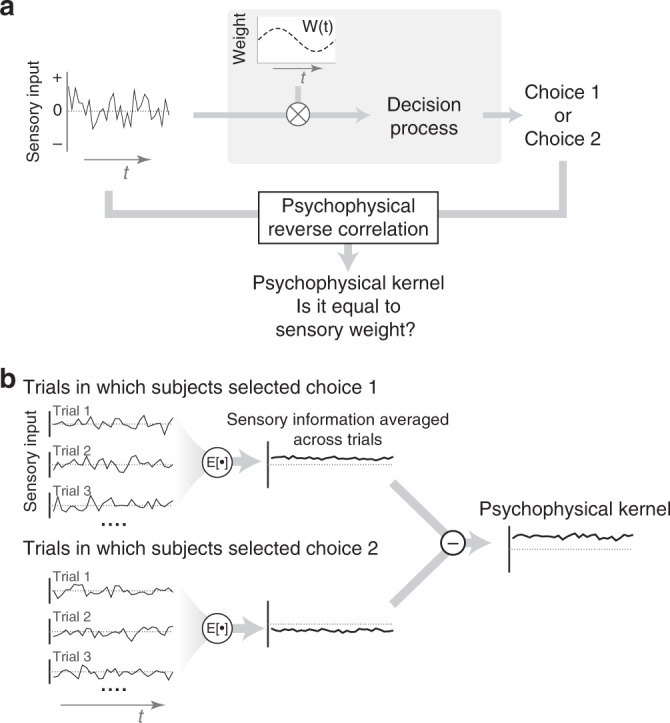
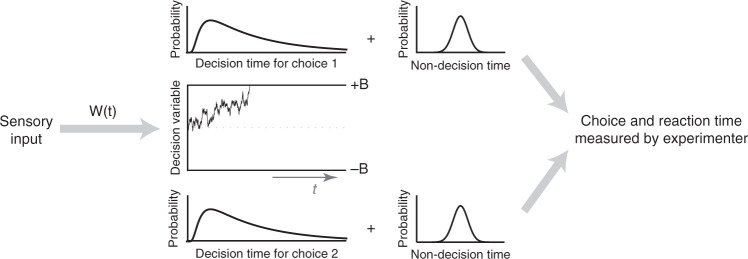
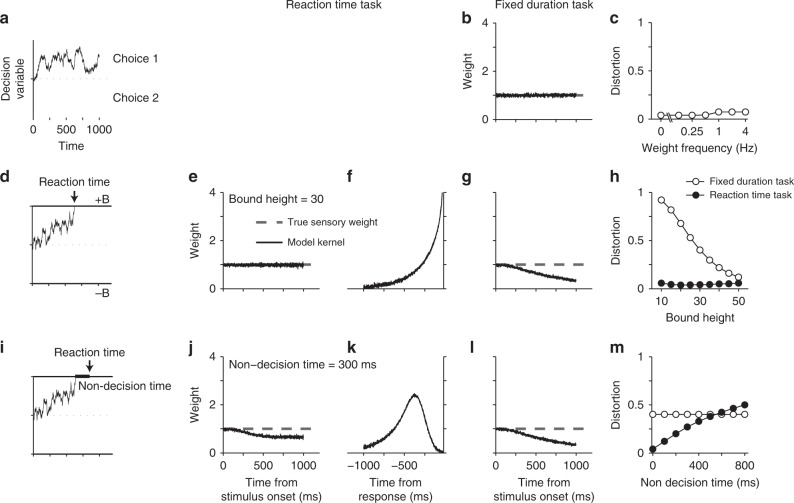
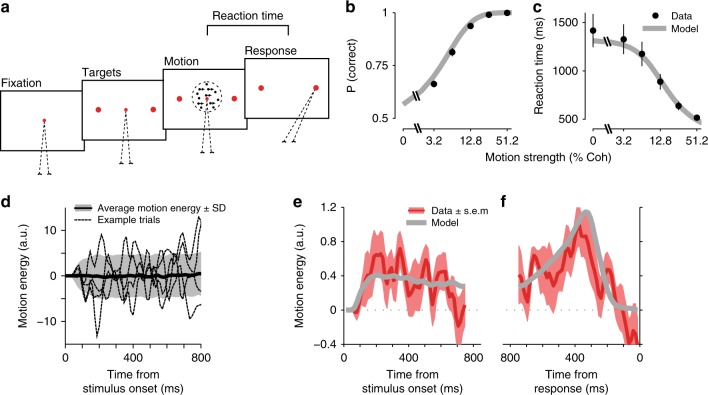
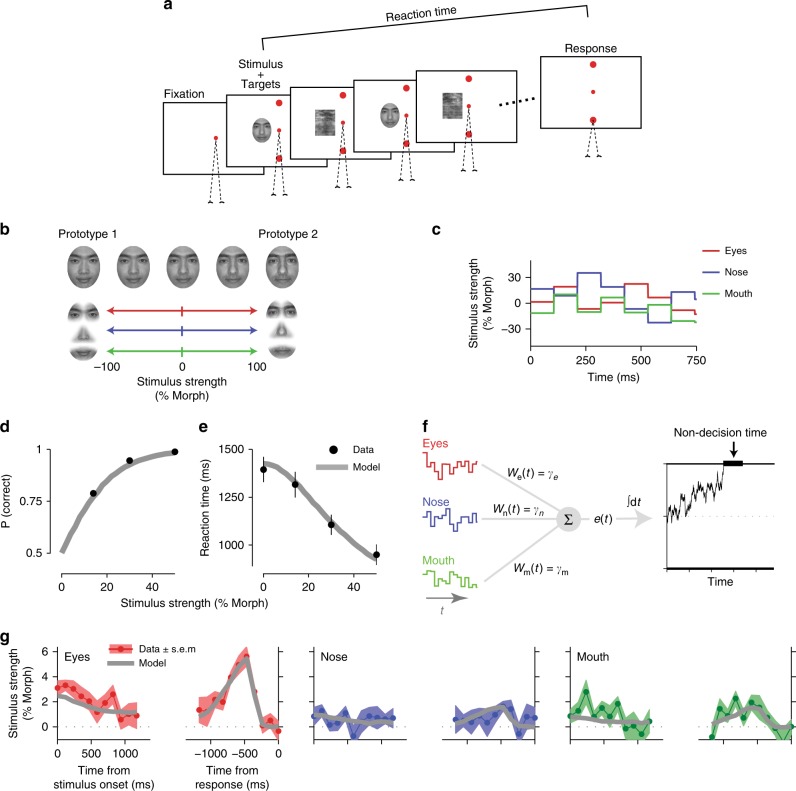
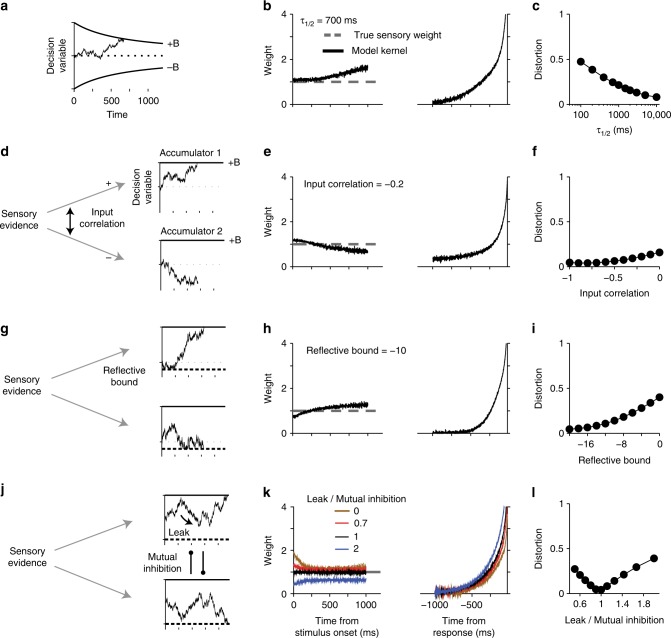
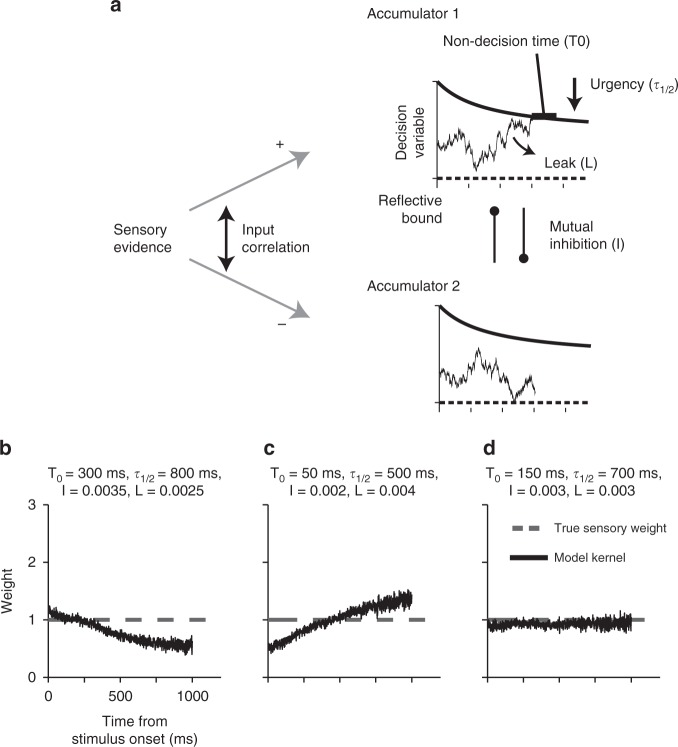
Goal-directed behavior depends on both sensory mechanisms that gather information from the outside world and decision-making mechanisms that select appropriate behavior based on that sensory information. Psychophysical reverse correlation is commonly used to quantify how fluctuations of sensory stimuli influence behavior and is generally believed to uncover the spatiotemporal weighting functions of sensory processes. Here we show that reverse correlations also reflect decision-making processes and can deviate significantly from the true sensory filters. Specifically, changes of decision bound and mechanisms of evidence integration systematically alter psychophysical reverse correlations. Similarly, trial-to-trial variability of sensory and motor delays and decision times causes systematic distortions in psychophysical kernels that should not be attributed to sensory mechanisms. We show that ignoring details of the decision-making process results in misinterpretation of reverse correlations, but proper use of these details turns reverse correlation into a powerful method for studying both sensory and decision-making mechanisms.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Ahumada Jr, A. J. Perceptual classification images from Vernier acuity masked by noise. Perception25, 1831–1840 (1996).
Publication types
MeSH terms
Grants and funding
- R90-1R90DA043849/U.S. Department of Health & Human Services | National Institutes of Health (NIH)/International
- P30 EY013079/EY/NEI NIH HHS/United States
- R90 DA043849/DA/NIDA NIH HHS/United States
- Sloan Research Fellowships/Alfred P. Sloan Foundation/International
- JSPS Postdoctoral Fellowship for Research Abroad/Japan Society for the Promotion of Science (JSPS)/International
- Simons Collaboration on the Global Brain/Simons Foundation/International
- McKnight Scholar Awards/McKnight Foundation/International
- Senior Fellowship in Biomedical Science/Charles H. Revson Foundation/International
- R01 MH109180/MH/NIMH NIH HHS/United States
- R01-MH109180/U.S. Department of Health & Human Services | National Institutes of Health (NIH)/International
LinkOut - more resources
Full Text Sources
Other Literature Sources
