

Intracellular C3 Protects Human Airway Epithelial Cells from Stress-associated Cell Death
- PMID: 30156437
- PMCID: PMC6376412
- DOI: 10.1165/rcmb.2017-0405OC
Intracellular C3 Protects Human Airway Epithelial Cells from Stress-associated Cell Death
Abstract
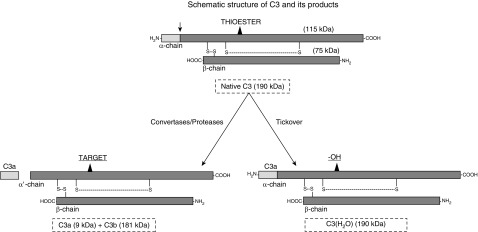
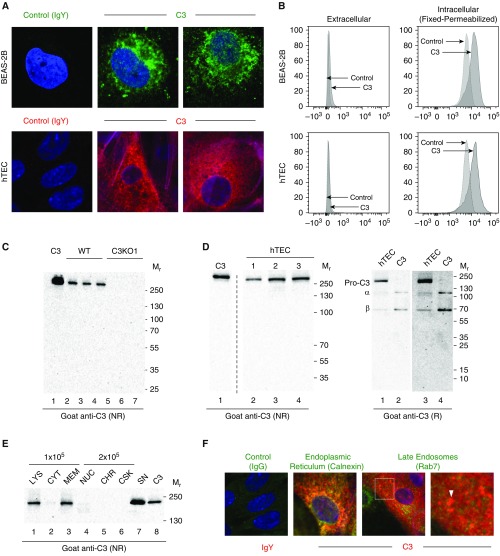
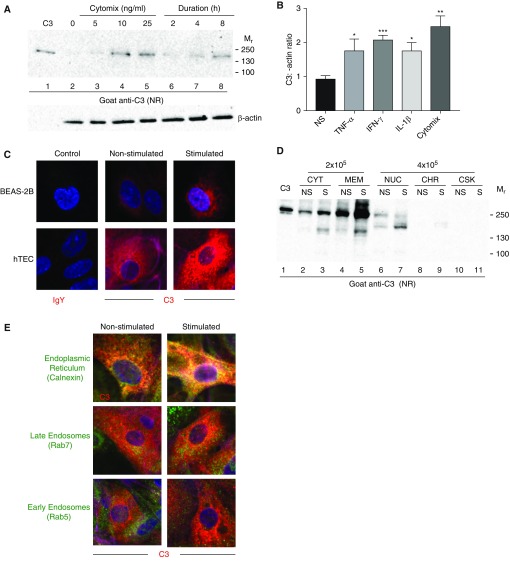
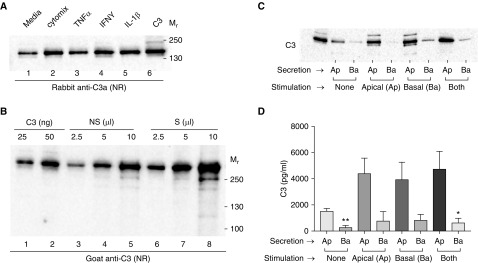
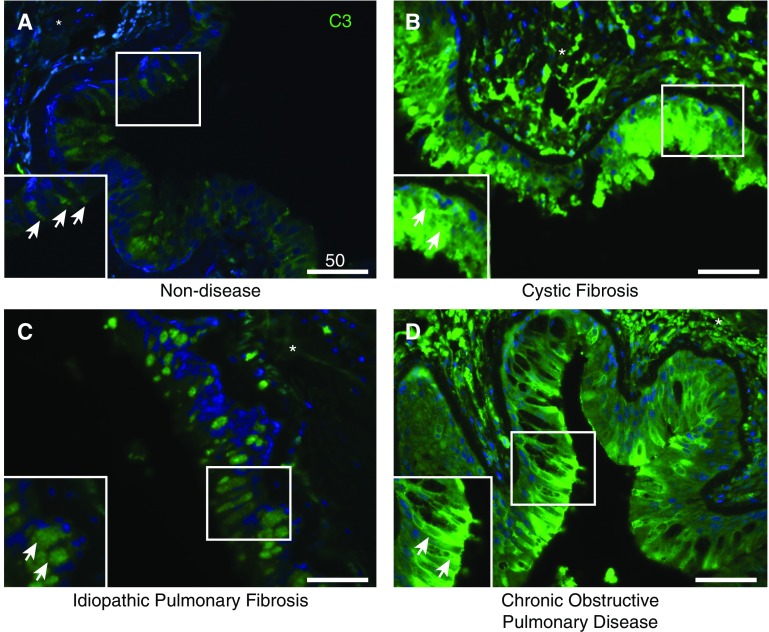
The complement system provides host defense against pathogens and environmental stress. C3, the central component of complement, is present in the blood and increases in BAL fluid after injury. We recently discovered that C3 is taken up by certain cell types and cleaved intracellularly to C3a and C3b. C3a is required for CD4+ T-cell survival. These observations made us question whether complement operates at environmental interfaces, particularly in the respiratory tract. We found that airway epithelial cells (AECs, represented by both primary human tracheobronchial cells and BEAS-2B [cell line]) cultured in C3-free media were unique from other cell types in that they contained large intracellular stores of de novo synthesized C3. A fraction of this protein reduced ("storage form") but the remainder did not, consistent with it being pro-C3 ("precursor form"). These two forms of intracellular C3 were absent in CRISPR knockout-induced C3-deficient AECs and decreased with the use of C3 siRNA, indicating endogenous generation. Proinflammatory cytokine exposure increased both stored and secreted forms of C3. Furthermore, AECs took up C3 from exogenous sources, which mitigated stress-associated cell death (e.g., from oxidative stress or starvation). C3 stores were notably increased within AECs in lung tissues from individuals with different end-stage lung diseases. Thus, at-risk cells furnish C3 through biosynthesis and/or uptake to increase locally available C3 during inflammation, while intracellularly, these stores protect against certain inducers of cell death. These results establish the relevance of intracellular C3 to airway epithelial biology and suggest novel pathways for complement-mediated host protection in the airway.
Keywords: anaphylatoxins; chronic obstructive pulmonary disease; cystic fibrosis; interstitial lung disease; oxidants.
Figures


References
-
- Circolo A, Garnier G, Fukuda W, Wang X, Hidvegi T, Szalai AJ, et al. Genetic disruption of the murine complement C3 promoter region generates deficient mice with extrahepatic expression of C3 mRNA. Immunopharmacology. 1999;42:135–149. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
