

Stat3 is indispensable for damage-induced crypt regeneration but not for Wnt-driven intestinal tumorigenesis
- PMID: 30156908
- PMCID: PMC6338624
- DOI: 10.1096/fj.201801176R
Stat3 is indispensable for damage-induced crypt regeneration but not for Wnt-driven intestinal tumorigenesis
Abstract
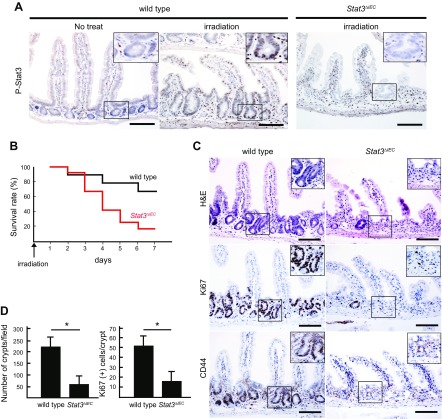
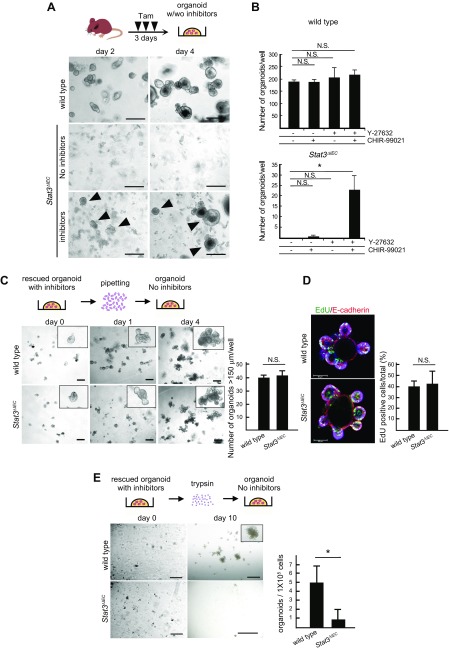
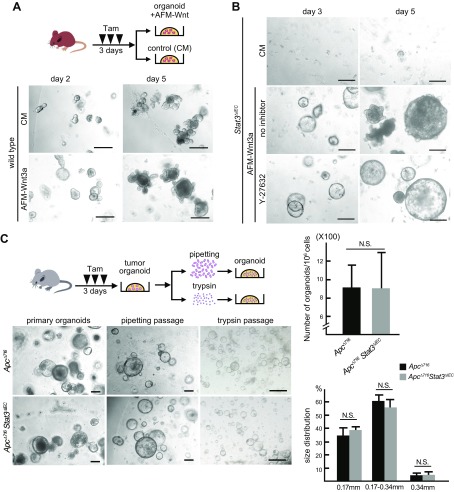
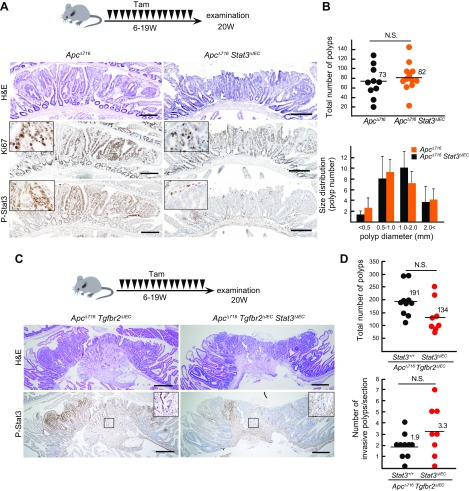
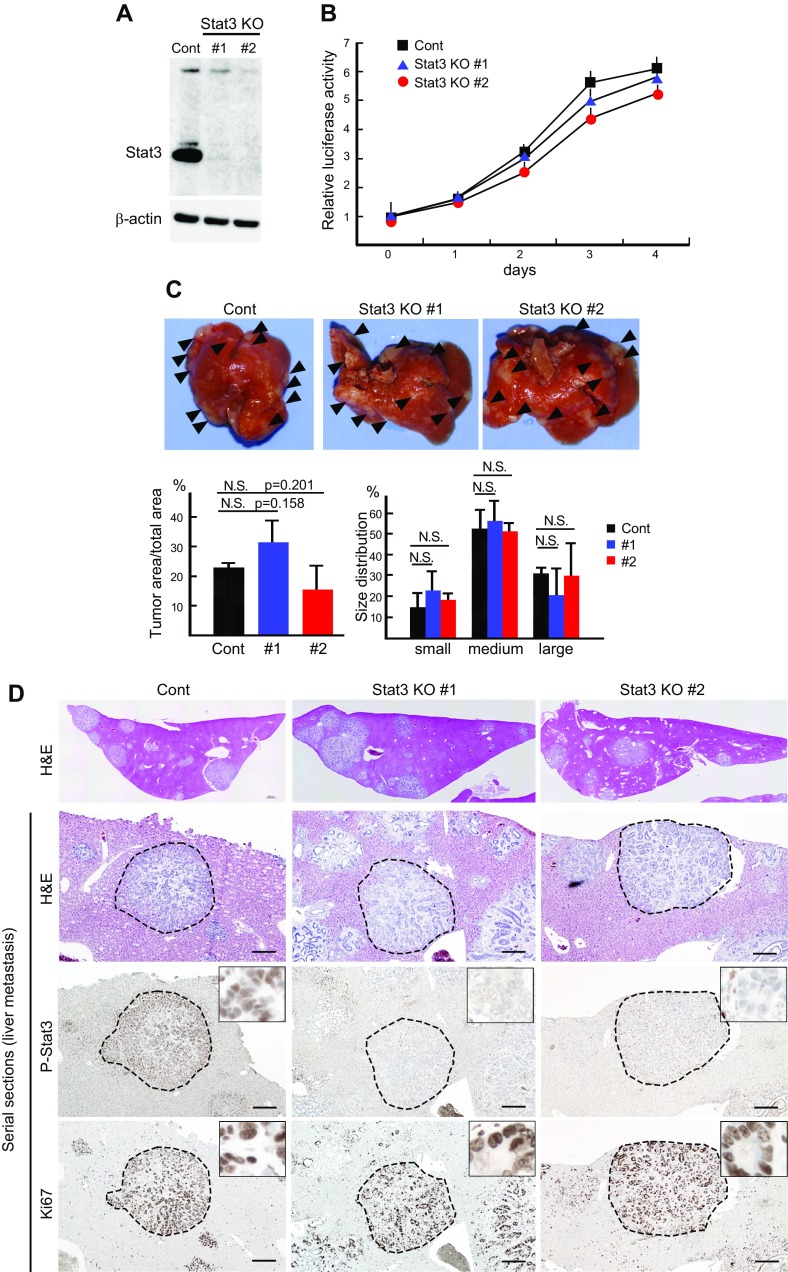
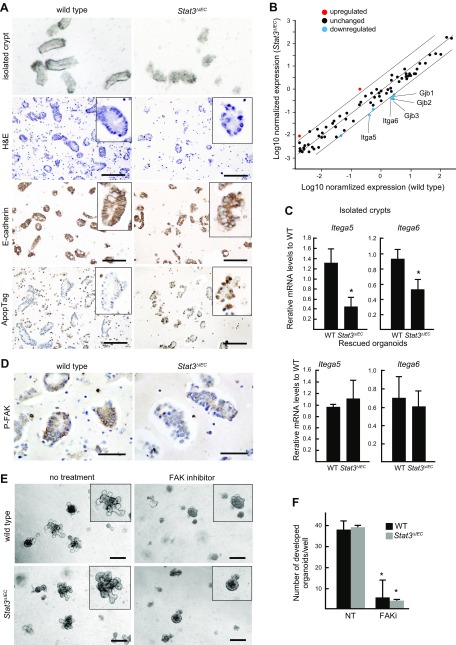
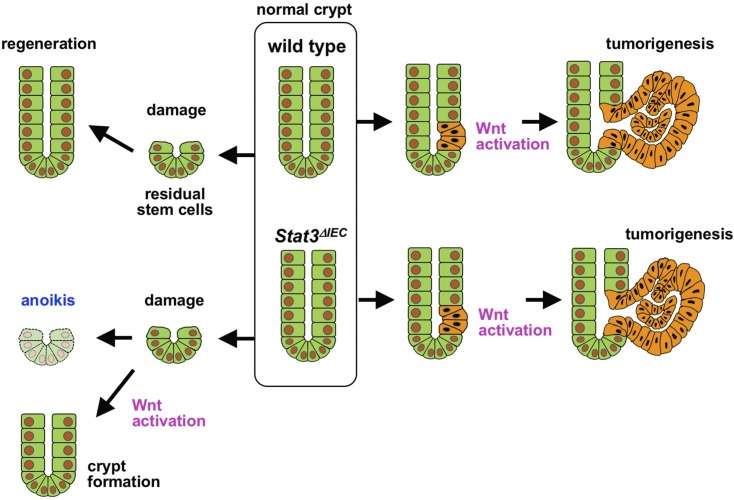
Signal transducer and activator of transcription 3 (Stat3) has been shown to play a role in intestinal regeneration and colitis-associated colon carcinogenesis. However, the role of Stat3 in the Wnt-driven sporadic intestinal tumorigenesis remains poorly understood. We examined the roles of Stat3 in intestinal regeneration and tumorigenesis by organoid culture experiments using Stat3∆IEC mouse-derived intestinal epithelial cells in which Stat3 was disrupted. The regeneration of intestinal mucosa and organoid formation were significantly suppressed by Stat3 disruption, which was compensated by Wnt activation. Furthermore, once organoids were recovered, Stat3 was no longer required for organoid growth. These results indicate that Stat3 and Wnt signaling cooperatively protect epithelial cells at the early phase of intestinal regeneration. In contrast, intestinal tumorigenesis was not suppressed by Stat3 disruption in adenomatous polyposis coli ( Apc) Δ716 and Apc∆716 Tgfbr2∆IEC mice, thus indicating that Stat3 is not required for Wnt activation-driven intestinal tumorigenesis. Mechanistically, Itga5 and Itga6 were down-regulated by Stat3 disruption, and focal adhesion kinase (FAK) activation was also suppressed. Notably, FAK inhibitor suppressed the organoid formation of wild-type epithelial cells. These results indicate that Stat3 is indispensable for the survival of epithelial cells through the activation of integrin signaling and the downstream FAK pathway; however, it is not required for the Wnt signaling-activated normal or tumor epithelial cells.-Oshima, H., Kok, S.-Y., Nakayama, M., Murakami, K., Voon, D. C.-C., Kimura, T., Oshima, M. Stat3 is indispensable for damage-induced crypt regeneration but not for Wnt-driven intestinal tumorigenesis.
Keywords: FAK; anoikis; colon cancer; integrin; organoids.
Conflict of interest statement
The authors thank M. Watanabe, A. Tsuda, and Y. Jomen (Division of Genetics, Cancer Research Institute, Kanazawa University) for their technical assistance. This study was supported by the Japan Agency for Medical Research and Development–Core Research for Evolutional Science and Technology (AMED–CREST) (JP17gm0410014) and AMED (JP17ck0106259); and Grants-in-Aid for Scientific Research (A) (JP18H04030) and (C) (JP16K07111) from the Ministry of Education, Culture, Sports, Science, and Technology of Japan; Takeda Science Foundation; and Mitsubishi Foundation. The authors declare no conflicts of interest.
Figures
References
-
- Frank, D. A. (2007) STAT3 as a central mediator of neoplastic cellular transformation. Cancer Lett. 251, 199–210 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
